

| Earthworm - Nervous System |




Above: the anterior part of the central nervous of the earthworm Lumbricus
terrestris. The nerve cord (shown in green) is ventral in position and runs from the
4th segment all the way to the last segment of the worm. A swelling or ganglion in
the cord occurs in each segment. These ganglia are small computers where nerve
cells communicate with each other, rather like regional brains, each responsible for
controlling its particular segment of the worm. Each of these ganglia puts out three
pairs of segmental nerves (i.e. there are 3 pairs per segment) though the medial
and posterior nerve roots join together just as they enter the nerve cord ganglion.
terrestris. The nerve cord (shown in green) is ventral in position and runs from the
4th segment all the way to the last segment of the worm. A swelling or ganglion in
the cord occurs in each segment. These ganglia are small computers where nerve
cells communicate with each other, rather like regional brains, each responsible for
controlling its particular segment of the worm. Each of these ganglia puts out three
pairs of segmental nerves (i.e. there are 3 pairs per segment) though the medial
and posterior nerve roots join together just as they enter the nerve cord ganglion.
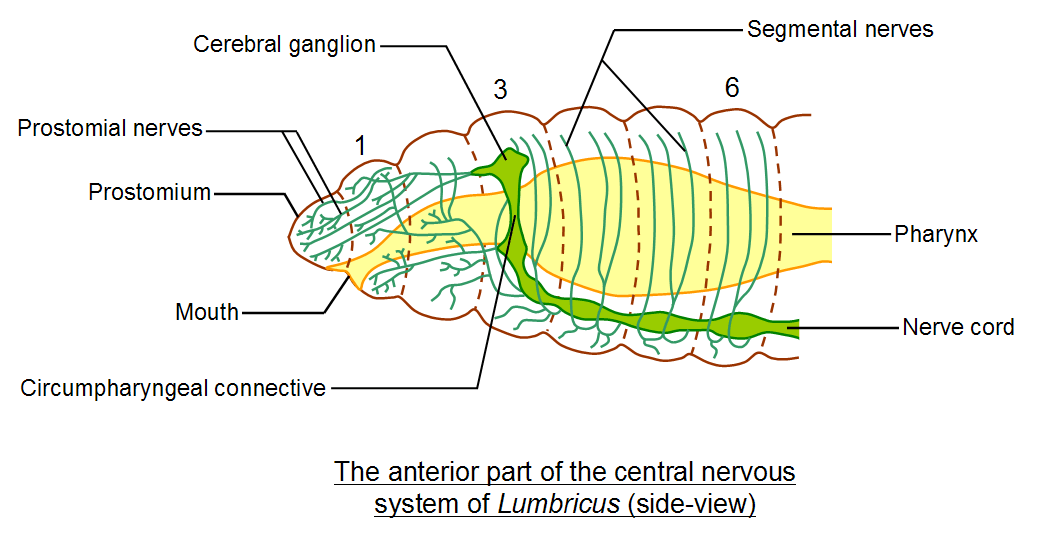
The ganglia of the first three segments are fused together to form a brain comprised of one pair of sub-pharyngeal ganglia
(ventral in position, beneath the pharynx of the gut) and one pair of cerebral ganglia (above the gut). One pair of
circumpharyngeal connectives (also called circumpharyngeal commissures, circumpharyngeal since the two connectives
surround the pharynx) connect the sub-pharyngeal and cerebral ganglia. Thus, the brain comprises a ganglionated ring
structure surrounding the fore-gut. The brain and ventral nerve cord together form the central nervous system (CNS). The
same structures are shown in plan view below:
(ventral in position, beneath the pharynx of the gut) and one pair of cerebral ganglia (above the gut). One pair of
circumpharyngeal connectives (also called circumpharyngeal commissures, circumpharyngeal since the two connectives
surround the pharynx) connect the sub-pharyngeal and cerebral ganglia. Thus, the brain comprises a ganglionated ring
structure surrounding the fore-gut. The brain and ventral nerve cord together form the central nervous system (CNS). The
same structures are shown in plan view below:
In the above diagram it can be seen that each circumpharyngeal connective gives off six short nerves to the gut. These nerves
then branch to form a gut plexus that runs between the circular and longitudinal muscle layers of the gut wall. Note that the
each of the posterior most pair of segmental nerves gives out a branch to innervate the intersegmental septum (these septa
are important in movement and locomotion). Each segmental nerve also consists of a ventral and a dorsal (lateral) branch.
The dorsal branch innervates the lateral and dorsal regions of the body wall, whilst the ventral branch innervates the ventral
region of the body wall. Each segmental nerve gives off many finer branches to innervate the muscles and sensors of the body
wall. The diagram below shows the pair of posterior segmental nerves in one segment seen end-on in cross-section:
then branch to form a gut plexus that runs between the circular and longitudinal muscle layers of the gut wall. Note that the
each of the posterior most pair of segmental nerves gives out a branch to innervate the intersegmental septum (these septa
are important in movement and locomotion). Each segmental nerve also consists of a ventral and a dorsal (lateral) branch.
The dorsal branch innervates the lateral and dorsal regions of the body wall, whilst the ventral branch innervates the ventral
region of the body wall. Each segmental nerve gives off many finer branches to innervate the muscles and sensors of the body
wall. The diagram below shows the pair of posterior segmental nerves in one segment seen end-on in cross-section:
The nerves that run along the body wall pass between the outer circular and inner longitudinal muscle layers of the wall. They
give off branches that form the intermuscular plexus and the subepidermal plexus. A plexus is essentially a web of nerve cells
connected together in a largely two dimensional grid or matrix. The subepidermal plexus occurs beneath the epidermis or outer
cell layer of the body wall and innervates the various sensors associated with the skin of the worm. The intermuscular plexus
penetrate the layers of muscle in the body wall and form chemical synapses with the muscle fibres which they control. There is a
separate sub-system for each of the longitudinal and circular muscle layers. Touching the worm will stimulate the subepidermal
nerve plexus which connects to the intermuscular plexus and causes the longitudinal muscles to contract. This reflex does not
require the CNS as it occurs even if the nerve cord is removed. Each segment, however, has its own nerve plexi and the plexus of
one segment is not connected directly to the nerve plexus of adjacent segments - the nerve cord is required to connect the
nervous systems of the segments together.
Scientists have ascertained many of these features by the following experiment:
They removed segments 40 to 60 from the worm, removed the gut and nerve cord from this portion and then turned the body wall
inside out and placed it in saline attached to levers that could record the pull of the muscles in the portion of body wall. Electrical
activity in the muscles and nerves could also be measured and the nervous system could be stimulated by a physical prod and by
electrodes. (These pieces of body wall even remained alive if kept in saline in the fridge overnight!).
When one segment of such a preparation is stimulated, the longitudinal muscles in that segment contracted and a wave of
contraction spread to the other segments, one after the other, but only when the segments were free to move, which showed that
it was simply the mechanical pull of the stimulated segment that stimulated the neighbouring segments to contract in turn.
Immobilising some of the segments stopped the spread of contraction, showing that the nerve plexus of one segment was not
connected to that of other segments. If the nerve cord was intact, however, then a nervous (electrical) signal passed along it to
the adjacent segments, even if some of the segments were immobilised. Thus, only the nerves in the nerve cord connect the
nervous systems of adjacent segments together.
give off branches that form the intermuscular plexus and the subepidermal plexus. A plexus is essentially a web of nerve cells
connected together in a largely two dimensional grid or matrix. The subepidermal plexus occurs beneath the epidermis or outer
cell layer of the body wall and innervates the various sensors associated with the skin of the worm. The intermuscular plexus
penetrate the layers of muscle in the body wall and form chemical synapses with the muscle fibres which they control. There is a
separate sub-system for each of the longitudinal and circular muscle layers. Touching the worm will stimulate the subepidermal
nerve plexus which connects to the intermuscular plexus and causes the longitudinal muscles to contract. This reflex does not
require the CNS as it occurs even if the nerve cord is removed. Each segment, however, has its own nerve plexi and the plexus of
one segment is not connected directly to the nerve plexus of adjacent segments - the nerve cord is required to connect the
nervous systems of the segments together.
Scientists have ascertained many of these features by the following experiment:
They removed segments 40 to 60 from the worm, removed the gut and nerve cord from this portion and then turned the body wall
inside out and placed it in saline attached to levers that could record the pull of the muscles in the portion of body wall. Electrical
activity in the muscles and nerves could also be measured and the nervous system could be stimulated by a physical prod and by
electrodes. (These pieces of body wall even remained alive if kept in saline in the fridge overnight!).
When one segment of such a preparation is stimulated, the longitudinal muscles in that segment contracted and a wave of
contraction spread to the other segments, one after the other, but only when the segments were free to move, which showed that
it was simply the mechanical pull of the stimulated segment that stimulated the neighbouring segments to contract in turn.
Immobilising some of the segments stopped the spread of contraction, showing that the nerve plexus of one segment was not
connected to that of other segments. If the nerve cord was intact, however, then a nervous (electrical) signal passed along it to
the adjacent segments, even if some of the segments were immobilised. Thus, only the nerves in the nerve cord connect the
nervous systems of adjacent segments together.
nerve cord is a double structure - it is really a pair of cords fused together. The cord is enclosed in a three-layered (trilaminar)
sheath. The outermost layer is a single layer of flat epithelial cells that are part of the peritoneum (the lining of the coelom).
Beneath this is a layer of longitudinal muscle cells, and beneath this is the innermost fibrous sheath that invests the nerve cells.
In animal nervous systems there are two principle types of nerve cell: the neurone which is the main signalling cell type,
frequently with many long processes or 'wires' projecting from it; and the neuroglia which are cells that form the insulation for
the neuronal 'wires' and also serve to keep the system clean, free from foreign bodies and perform various other maintenance
and protective functions - protecting the delicate wiring of the nervous system. The main wires in the system are long slender
processes from neurones, and these processes are called axons. There are many axons in the nerve cord, but most are small,
however, five are especially large and easily seen under the microscope - these are the giant axons. These are axons with very
large diameters. The dorsal giant axons is the widest, with a diameter of about 50 micrometres. An axon is rather like an
electrical wire, and just as electricity flows more easily and faster along a wide wire than along a narrow wire (which provides
more resistance to the flow of electricity) so the nervous signal flows faster along giant axons. The nervous signal is positive
electricity, consisting of the flow of positively charged ions, whereas electricity in electrical wires is negative electricity, consisting
of the flow of negatively charged electrons.
These giant axons are used to carry the fastest signals along the nerve cord, and these are emergency signals that initiate
escape reflexes. The larger dorsal giant axon conducts signals the fastest and in the direction from aft to fore only. This sends
signals from the rear of the worm forwards, such that if something unexpected touches the rear of the worm, a rapid signal is
sent forwards along the nerve cord, causing the longitudinal muscle sin each segment to rapidly contract, shortening the worm
rapidly in an attempt to escape from a would-be predator or other threat. The two dorso-lateral giant axons connect with each
other and send signals from the front end to the rear, in case the worm has to make a rapid retreat (perhaps contracting into its
burrow to escape a curious bird).
The giant axons are made up of a number of cylindrical cells all connected end-to-end. The junctions between neighbouring
cells contain fast electrical synapses (nexuses) that allow the electrical signal to rapidly travel from one cell to another on its way
down the axon.
Sensory systems
The CNS is a series of tiny (but powerful!) computers that form the brain and ventral nerve cord of the worm. This sends out
nerves, such as the segmental nerves, to the muscles and sensors of the worm. The job of the CNS is to analyse the incoming
sensory information from the various sensors and determine a course of action and then send out the appropriate instructions
to the muscles.
One type of important sensor are photoreceptors (light sensors). Earthworms have no eyes (although many other worms do,
eyes are not so useful when you spend most of your life buried in soil or crawling about at night). However, worms do have light
sensors in the form of specialised nerve endings (called light cells of Hess). These sensors occur in most parts of the worm's
skin, but they are concentrated in certain areas. They are more concentrated on the back and sides of the worm, though a few
do occur on the ventral surface at the front end (1st segment) of the worm. They are also much more concentrated toward the
front end of the worm, being most numerous in the prostomium (the front-most lobe which is not a true segment) and dwindling
in density in the first three segments and are very few in number past the third segment. There are also quite a few on the
rear-most segment of the worm, including its ventral surface. (Note that the front and back ends of the worm are often lifted up
above the ground and so having some light sensors underneath the front and back ends, as well as one the sides and back,
makes sense).
So, the prostomium is the main light-sensing region of the worm. The prostomium is innervated by a pair of branching nerves
from the cerebral ganglia. These branches from the subepidermal and intermuscular plexuses. Some of the fine branches in the
subepidermal plexus end in photoreceptors (light-sensitive cells). Additionally, many of the larger branches have bulbous
clusters of photoreceptors part-way along their length inside the prostomium coelom. Remember that the prostomium is not very
large - it is the pointed 'nose' of the worm, forming the upper lip that overhangs the mouth and so some light can shine all the
way through it in bright sunlight. Presumably by comparing the signals coming from the subepidermal photoreceptors wwith
those coming from the more shaded photoreceptors deeper within the prostomium the worm can better gauge the light intensity.
The three thumbnails below illustrate the arrangement of these photoreceptors within the prostomium (click to enlarge):
sheath. The outermost layer is a single layer of flat epithelial cells that are part of the peritoneum (the lining of the coelom).
Beneath this is a layer of longitudinal muscle cells, and beneath this is the innermost fibrous sheath that invests the nerve cells.
In animal nervous systems there are two principle types of nerve cell: the neurone which is the main signalling cell type,
frequently with many long processes or 'wires' projecting from it; and the neuroglia which are cells that form the insulation for
the neuronal 'wires' and also serve to keep the system clean, free from foreign bodies and perform various other maintenance
and protective functions - protecting the delicate wiring of the nervous system. The main wires in the system are long slender
processes from neurones, and these processes are called axons. There are many axons in the nerve cord, but most are small,
however, five are especially large and easily seen under the microscope - these are the giant axons. These are axons with very
large diameters. The dorsal giant axons is the widest, with a diameter of about 50 micrometres. An axon is rather like an
electrical wire, and just as electricity flows more easily and faster along a wide wire than along a narrow wire (which provides
more resistance to the flow of electricity) so the nervous signal flows faster along giant axons. The nervous signal is positive
electricity, consisting of the flow of positively charged ions, whereas electricity in electrical wires is negative electricity, consisting
of the flow of negatively charged electrons.
These giant axons are used to carry the fastest signals along the nerve cord, and these are emergency signals that initiate
escape reflexes. The larger dorsal giant axon conducts signals the fastest and in the direction from aft to fore only. This sends
signals from the rear of the worm forwards, such that if something unexpected touches the rear of the worm, a rapid signal is
sent forwards along the nerve cord, causing the longitudinal muscle sin each segment to rapidly contract, shortening the worm
rapidly in an attempt to escape from a would-be predator or other threat. The two dorso-lateral giant axons connect with each
other and send signals from the front end to the rear, in case the worm has to make a rapid retreat (perhaps contracting into its
burrow to escape a curious bird).
The giant axons are made up of a number of cylindrical cells all connected end-to-end. The junctions between neighbouring
cells contain fast electrical synapses (nexuses) that allow the electrical signal to rapidly travel from one cell to another on its way
down the axon.
Sensory systems
The CNS is a series of tiny (but powerful!) computers that form the brain and ventral nerve cord of the worm. This sends out
nerves, such as the segmental nerves, to the muscles and sensors of the worm. The job of the CNS is to analyse the incoming
sensory information from the various sensors and determine a course of action and then send out the appropriate instructions
to the muscles.
One type of important sensor are photoreceptors (light sensors). Earthworms have no eyes (although many other worms do,
eyes are not so useful when you spend most of your life buried in soil or crawling about at night). However, worms do have light
sensors in the form of specialised nerve endings (called light cells of Hess). These sensors occur in most parts of the worm's
skin, but they are concentrated in certain areas. They are more concentrated on the back and sides of the worm, though a few
do occur on the ventral surface at the front end (1st segment) of the worm. They are also much more concentrated toward the
front end of the worm, being most numerous in the prostomium (the front-most lobe which is not a true segment) and dwindling
in density in the first three segments and are very few in number past the third segment. There are also quite a few on the
rear-most segment of the worm, including its ventral surface. (Note that the front and back ends of the worm are often lifted up
above the ground and so having some light sensors underneath the front and back ends, as well as one the sides and back,
makes sense).
So, the prostomium is the main light-sensing region of the worm. The prostomium is innervated by a pair of branching nerves
from the cerebral ganglia. These branches from the subepidermal and intermuscular plexuses. Some of the fine branches in the
subepidermal plexus end in photoreceptors (light-sensitive cells). Additionally, many of the larger branches have bulbous
clusters of photoreceptors part-way along their length inside the prostomium coelom. Remember that the prostomium is not very
large - it is the pointed 'nose' of the worm, forming the upper lip that overhangs the mouth and so some light can shine all the
way through it in bright sunlight. Presumably by comparing the signals coming from the subepidermal photoreceptors wwith
those coming from the more shaded photoreceptors deeper within the prostomium the worm can better gauge the light intensity.
The three thumbnails below illustrate the arrangement of these photoreceptors within the prostomium (click to enlarge):
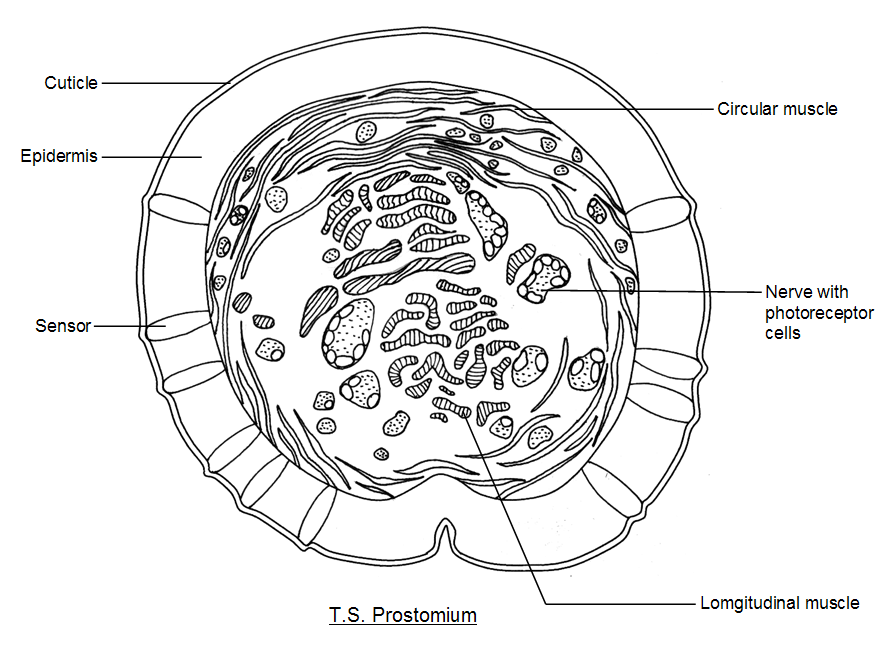
Above: The prostomium of Lumbricus terrestris. left, a longitudinal section through the prostomium, showing the prostomial
photoreceptors (nearer to the surface) and the deeper clusters of photoreceptors and the sub-epidermal nerve plexus. Also
shown are sensors in the epidermis consisting of ellipsoidal clusters of elongated epidermal cells (epidermal sensors) bearing
cilia (projections, possibly immotile sensory cilia) on their external apex. These sensors are found scattered around the
earthworm and may be touch sensors or chemoreceptors. Right, a cross-section (transverse section) through the prostomium
showing the photoreceptors, nerves and muscle cells. Notice how the epidermal sensor units are strategically placed along the
sides of the prostomium.
The photoreceptors of Lumbricus are about as simple as photoreceptors get - they are the specialised endings of nerve cells.
All living cells appear capable of responding to light, so nerve cells should all be capable of detecting light, in principle.
However, the light cells of Hess are specially modified to make them especially sensitive to light. Much depends of course on
how the CNS analyses the incoming data from the sensory nerves around the body. Weak signals must be filtered out to
remove background noise and it is doubtful that ordinary nerve cells are involved in light perception.
No doubt much remains to be learned about earthworm sensory systems - the whole subject seems very poorly studied to me,
though I shall continue to research the literature in this area and bring you any important updates.
photoreceptors (nearer to the surface) and the deeper clusters of photoreceptors and the sub-epidermal nerve plexus. Also
shown are sensors in the epidermis consisting of ellipsoidal clusters of elongated epidermal cells (epidermal sensors) bearing
cilia (projections, possibly immotile sensory cilia) on their external apex. These sensors are found scattered around the
earthworm and may be touch sensors or chemoreceptors. Right, a cross-section (transverse section) through the prostomium
showing the photoreceptors, nerves and muscle cells. Notice how the epidermal sensor units are strategically placed along the
sides of the prostomium.
The photoreceptors of Lumbricus are about as simple as photoreceptors get - they are the specialised endings of nerve cells.
All living cells appear capable of responding to light, so nerve cells should all be capable of detecting light, in principle.
However, the light cells of Hess are specially modified to make them especially sensitive to light. Much depends of course on
how the CNS analyses the incoming data from the sensory nerves around the body. Weak signals must be filtered out to
remove background noise and it is doubtful that ordinary nerve cells are involved in light perception.
No doubt much remains to be learned about earthworm sensory systems - the whole subject seems very poorly studied to me,
though I shall continue to research the literature in this area and bring you any important updates.
Structure of the body wall and respiration Movement, support and locomotion Circulatory system Nervous and sensory systems Nutrition Excretion Reproduction and development Ecology Worm evolution |
| | | | | | | | |
| | | | | | | | | |