
| |
|
|
|
|
|
|
|










| Transport
in Plants |
Carbon
Fixation by Roots
The main source of carbon for plants comes from the atmosphere as carbon dioxide and fixation of this carbon dioxide by leaves and other
green parts of the shoot system in the presence of light as an energy source - the process of photosynthesis. Some of this fixed carbon is
converted into fuels for respiration, such as glucose, some is stored as compact materials like starch, and some is used in building blocks,
such as proteins, nucelotides and phospholipids, to build the plant body.
Fixation is an odd word, and one that stems all the way back to alchemy. It makes perfect sense, however, when you think of gases as
mobile, with their molecules constantly moving around by diffusion due to their thermal kinetic energy. Fixation means to render less
mobile, and was historically depicted by clipping the 'wings' of the gaseous or volatile substance (by converting it into a solid or
non-volatile liquid, for example).
However, it is a little known fact that roots can also fix carbon. Roots do not photosynthesise, however, and they fix carbon in the absence
of light with the use of enzymes. Ordinarily roots only fix enough carbon for their own secretory activities, with the bulk of carbon fixation
occurring in the shoot system. Root-fixed carbon is used, for example, to produce root secretions which leak into the soil around the roots
(the rhizosphere). These secretions probably have various functions, but appear to encourage plant-friendly microbes, signaling to
mycorhizal fungi, for example, advertising the root and encouraging the fungus to form a symbiosis with it. These secretions are also
thought to be important in helping to 'mobilise, plant nutrients - many nutrients are locked onto soil particles in an insoluble form and so
can not be absorbed by the roots until they have been dissolved or mobilised. In some herbaceous plants, it has been shown that roots
can fix enough surplus carbon to contribute to shoot carbon, that is the roots export carbon that they fix and which is in excess of their own
needs, into the shoot system. To what extent the roots contribute to the stem carbon of woody plants is not known.
The Role of Root Pressure
As well as contributing a small amount to the pressure gradient which drives xylem sap part-way up a tree, or perhaps to the top of some
short plants, or even small trees, in certain conditions, root pressure possibly has several important functions. In very humid climates
where evapotranspiration is too low to drive xylem flow, root pressure ensures that some xylem travels up the plant to deliver mineral
nutrients from the roots to the shoot system. This root pressure may cause droplets of water to exude from vessels at the edges and tips
of leaves (guttation) and some plants have special pores (hydathodes) at the ends of the veins to allow this water out (after the minerals
are extracted from it by the cells). This flow due to root pressure also occurs in many plants at night, contributing to early morning due. It is
also important in very early spring, supplying growing buds with minerals before the leaves have opened enough for evapotranspiration to
take over as the main driving force. Root pressure may also help unblock cavitated vessels.
Sugar in the Xylem
In maples and birches, cold-tolerant trees, xylem sap is driven up the stem in winter and carries sucrose with it to fuel the developing
flowers which open early before the leaves. The sucrose is loaded into the xylem from ray parenchyma and other storage cells in the
xylem. This sap ascent can not be utilising the transpiration stream as no leaves are available in winter to drive it. It occurs on warmer
days that follow cold nights and is thought to involve a night-freeze, warm-day cycle of pressure changes in the trunk. At night the xylem
sap freezes and this is thought to trap and compress gases in the xylem as the sap freezes. The daytime heat melts the ice in the xylem,
expanding the trapped gases and generating a positive pressure to drive xylem sap flow up the trunk. At this point the sugary xylem sap
can be collected for use in making syrup and wines. In the evenings, the colling temperatures dissolve more gases in the xylem, seeded
from adjoining tissues such as parenchyma, ready to form compressed gases when the xylem freezes at night. A good article, on an
external web site, describing this process in maple can be found here: maple syrup.
Foliar Absorption
It should be remembered that plants can also absorb nutrients through their leaves. Gases, liquids and solid mineral dusts falling on the
leaf can all lead to increased nutrient absorption. The nutrients are absorbed through the plasmodesmata and also to some extent
through the cuticle. Apparently, a European forest may absorb up to 30% of its nitrogen through the canopy in this way (Thomas, 2000).
Bibliography
Schulz, 1992. Living sieve cells of conifers as visualized by confocal, laser-scanning fluorescence microscopy. Protoplasma 166: 153-164.
Scheirer, 1978. Cell wall chemistry and fine structure in leptoids of Dendroligotrichum (Bryophyta): The end wall. Amer. J. Bot. 65:
1027-1031.
Esau, 1976. Anatomy of Seed plants, second edition.
Thomas, P. 2000. Trees: their natural history. Cambridge University Press.
Other sources to be added...
Article last updated: 27/2/15, 25 Aug 2020 (some broken links fixed)
The main source of carbon for plants comes from the atmosphere as carbon dioxide and fixation of this carbon dioxide by leaves and other
green parts of the shoot system in the presence of light as an energy source - the process of photosynthesis. Some of this fixed carbon is
converted into fuels for respiration, such as glucose, some is stored as compact materials like starch, and some is used in building blocks,
such as proteins, nucelotides and phospholipids, to build the plant body.
Fixation is an odd word, and one that stems all the way back to alchemy. It makes perfect sense, however, when you think of gases as
mobile, with their molecules constantly moving around by diffusion due to their thermal kinetic energy. Fixation means to render less
mobile, and was historically depicted by clipping the 'wings' of the gaseous or volatile substance (by converting it into a solid or
non-volatile liquid, for example).
However, it is a little known fact that roots can also fix carbon. Roots do not photosynthesise, however, and they fix carbon in the absence
of light with the use of enzymes. Ordinarily roots only fix enough carbon for their own secretory activities, with the bulk of carbon fixation
occurring in the shoot system. Root-fixed carbon is used, for example, to produce root secretions which leak into the soil around the roots
(the rhizosphere). These secretions probably have various functions, but appear to encourage plant-friendly microbes, signaling to
mycorhizal fungi, for example, advertising the root and encouraging the fungus to form a symbiosis with it. These secretions are also
thought to be important in helping to 'mobilise, plant nutrients - many nutrients are locked onto soil particles in an insoluble form and so
can not be absorbed by the roots until they have been dissolved or mobilised. In some herbaceous plants, it has been shown that roots
can fix enough surplus carbon to contribute to shoot carbon, that is the roots export carbon that they fix and which is in excess of their own
needs, into the shoot system. To what extent the roots contribute to the stem carbon of woody plants is not known.
The Role of Root Pressure
As well as contributing a small amount to the pressure gradient which drives xylem sap part-way up a tree, or perhaps to the top of some
short plants, or even small trees, in certain conditions, root pressure possibly has several important functions. In very humid climates
where evapotranspiration is too low to drive xylem flow, root pressure ensures that some xylem travels up the plant to deliver mineral
nutrients from the roots to the shoot system. This root pressure may cause droplets of water to exude from vessels at the edges and tips
of leaves (guttation) and some plants have special pores (hydathodes) at the ends of the veins to allow this water out (after the minerals
are extracted from it by the cells). This flow due to root pressure also occurs in many plants at night, contributing to early morning due. It is
also important in very early spring, supplying growing buds with minerals before the leaves have opened enough for evapotranspiration to
take over as the main driving force. Root pressure may also help unblock cavitated vessels.
Sugar in the Xylem
In maples and birches, cold-tolerant trees, xylem sap is driven up the stem in winter and carries sucrose with it to fuel the developing
flowers which open early before the leaves. The sucrose is loaded into the xylem from ray parenchyma and other storage cells in the
xylem. This sap ascent can not be utilising the transpiration stream as no leaves are available in winter to drive it. It occurs on warmer
days that follow cold nights and is thought to involve a night-freeze, warm-day cycle of pressure changes in the trunk. At night the xylem
sap freezes and this is thought to trap and compress gases in the xylem as the sap freezes. The daytime heat melts the ice in the xylem,
expanding the trapped gases and generating a positive pressure to drive xylem sap flow up the trunk. At this point the sugary xylem sap
can be collected for use in making syrup and wines. In the evenings, the colling temperatures dissolve more gases in the xylem, seeded
from adjoining tissues such as parenchyma, ready to form compressed gases when the xylem freezes at night. A good article, on an
external web site, describing this process in maple can be found here: maple syrup.
Foliar Absorption
It should be remembered that plants can also absorb nutrients through their leaves. Gases, liquids and solid mineral dusts falling on the
leaf can all lead to increased nutrient absorption. The nutrients are absorbed through the plasmodesmata and also to some extent
through the cuticle. Apparently, a European forest may absorb up to 30% of its nitrogen through the canopy in this way (Thomas, 2000).
Bibliography
Schulz, 1992. Living sieve cells of conifers as visualized by confocal, laser-scanning fluorescence microscopy. Protoplasma 166: 153-164.
Scheirer, 1978. Cell wall chemistry and fine structure in leptoids of Dendroligotrichum (Bryophyta): The end wall. Amer. J. Bot. 65:
1027-1031.
Esau, 1976. Anatomy of Seed plants, second edition.
Thomas, P. 2000. Trees: their natural history. Cambridge University Press.
Other sources to be added...
Article last updated: 27/2/15, 25 Aug 2020 (some broken links fixed)
We've
all sat under enormous old trees, whether it's been for a picnic,
or for shade, or just to have a quiet place to sit and
think. Many of us vaguely remember our school science classes and can recall the basics of photosynthesis and the life-cycle
of a tree. However, there's actually a lot more going on under that bark than you might realize. Take another look at how a tree
waters, feeds and grows and all the intricate mechanisms it has developed to do it and you may find it quite fascinating! At the
least, it'll certainly give you something to consider the next time you lean up against a 100 year old oak tree to consider your
finances.
Oak trees, like the one illustrated above (a 3D Pov-Ray model) may take-up more than 400 litres of water each day! This water
moves up the stem in the outermost rings of xylem (wood). Some of this water becomes (temporarily) incorporated into the cells
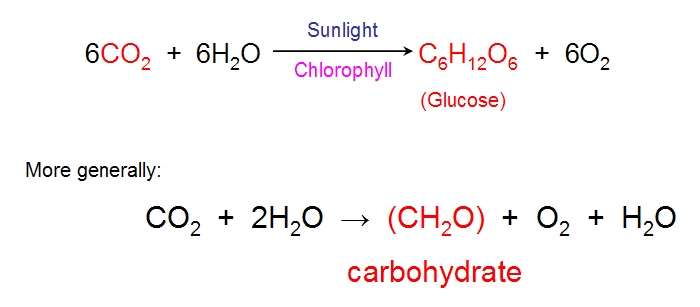
have a vast surface area of leaves to catch enough sunlight for photosynthesis and these leaves need carbon dioxide which is
reacted with water to form the organic building blocks of the plant's cells, using the energy harnessed by sunlight. These
building blocks include amino acids and sugars. The much needed carbon dioxide comes from the atmosphere and leaves
have closable pores called stomata (singular stoma) to take up this carbon dioxide by diffusion. Leaves are generally born on
stalks which have a hinge at their base (called the pulvinus) and this allows the leaves to vibrate in the wind, stirring the air to
help the leaves obtain a fresh supply of carbon dioxide (diffusion in still air is slow and a zone of stagnant carbon dioxide
depleted air would surround each leaf). A general equation for photosynthesis is given below:
think. Many of us vaguely remember our school science classes and can recall the basics of photosynthesis and the life-cycle
of a tree. However, there's actually a lot more going on under that bark than you might realize. Take another look at how a tree
waters, feeds and grows and all the intricate mechanisms it has developed to do it and you may find it quite fascinating! At the
least, it'll certainly give you something to consider the next time you lean up against a 100 year old oak tree to consider your
finances.
Oak trees, like the one illustrated above (a 3D Pov-Ray model) may take-up more than 400 litres of water each day! This water
moves up the stem in the outermost rings of xylem (wood). Some of this water becomes (temporarily) incorporated into the cells
have a vast surface area of leaves to catch enough sunlight for photosynthesis and these leaves need carbon dioxide which is
reacted with water to form the organic building blocks of the plant's cells, using the energy harnessed by sunlight. These
building blocks include amino acids and sugars. The much needed carbon dioxide comes from the atmosphere and leaves
have closable pores called stomata (singular stoma) to take up this carbon dioxide by diffusion. Leaves are generally born on
stalks which have a hinge at their base (called the pulvinus) and this allows the leaves to vibrate in the wind, stirring the air to
help the leaves obtain a fresh supply of carbon dioxide (diffusion in still air is slow and a zone of stagnant carbon dioxide
depleted air would surround each leaf). A general equation for photosynthesis is given below:


Left: a
section through a leaf blade. Both
the upper and lower surfaces of the leaf
are bounded by a layer of epithelium (E)
(covering tissue) which is coated by a
cuticle (C) of its own secretion. The
cuticle is thickest on the upper surface of
the leaf and contains waxy materials to
make it water proof in order to reduce
water loss by evaporation. Stomata occur
mostly among the lower epidermis and
each stoma (S) is surrounded by a pair
of sausage-shaped guard cells (red).
the upper and lower surfaces of the leaf
are bounded by a layer of epithelium (E)
(covering tissue) which is coated by a
cuticle (C) of its own secretion. The
cuticle is thickest on the upper surface of
the leaf and contains waxy materials to
make it water proof in order to reduce
water loss by evaporation. Stomata occur
mostly among the lower epidermis and
each stoma (S) is surrounded by a pair
of sausage-shaped guard cells (red).
(palisade mesophyll, PM) whilst the lower
leaf contains more
loosely packed isodiametric cells (spongy mesophyll, SM).
traversing the middle of the leaf blade are vessels which branch
from the central midrib. Each vessel or vascular bundle is
enclosed in a cylinder of cells called the bundle sheath (BS) and
contains two principle types of vascular or water-conducting
tissue: xylem (X) and phloem (P). Note the many spaces
between the cells, especially the spongy mesophyll - these are a
series of interconnected air spaces. The airs spaces connect
to the outside air through the stomata via an air space beneath
each stoma, called the sub-stomatal air space (visible in this
photomicrograph). Air carrying carbon dioxide diffuses into the
leaf through the stomata and then diffuses throughout the
air-space chambers to reach the mesophyll. The mesophyll
(especially the palisade mesophyll) are the site of
photosynthesis and contain chloroplasts which contain the
green light-harvesting pigment chlorophyll. In plants, epidermal
cells do not contain chloroplasts (with the exception of guard
cells) and so do not photosynthesise.
loosely packed isodiametric cells (spongy mesophyll, SM).
traversing the middle of the leaf blade are vessels which branch
from the central midrib. Each vessel or vascular bundle is
enclosed in a cylinder of cells called the bundle sheath (BS) and
contains two principle types of vascular or water-conducting
tissue: xylem (X) and phloem (P). Note the many spaces
between the cells, especially the spongy mesophyll - these are a
series of interconnected air spaces. The airs spaces connect
to the outside air through the stomata via an air space beneath
each stoma, called the sub-stomatal air space (visible in this
photomicrograph). Air carrying carbon dioxide diffuses into the
leaf through the stomata and then diffuses throughout the
air-space chambers to reach the mesophyll. The mesophyll
(especially the palisade mesophyll) are the site of
photosynthesis and contain chloroplasts which contain the
green light-harvesting pigment chlorophyll. In plants, epidermal
cells do not contain chloroplasts (with the exception of guard
cells) and so do not photosynthesise.

Left: a
varnish cast of the undersurface of a leaf showing the
stomata and their accompanying guard cells.
Click here to see more about leaf morphology and vasculature.
stomata and their accompanying guard cells.
Click here to see more about leaf morphology and vasculature.
Stomata
will close to conserve water at night-time when photosynthesis can
no longer continue, and they will close if the plant is
losing too much water. Stomata will open and close according to a circadian rhythm as well as in direct response to light and dark.
Stomata may also close in response to wounding - plants can lose a lot of water through open wounds and some plants, e.g.
tomato plants, react rapidly to damage by transmitting electrical signals throughout their leaves which trigger the stomata to close.
Despite these measures, plants lose a lot of water through their stomata and this water needs to be replaced. Xylem vessels carry
water from the roots, up the stem and to the leaves. Plants exploit this situation, making the most of a 'bad thing' and they utilise
the so-called transpiration stream of sap ascending in the xylem to carry mineral nutrients, obtained by the roots from the soil,
with it. Transpiration from the leaves also helps to keep them cool (important as they are purposefully exposed to the sunlight!).
The xylem sap is carried into each leaf, along the central vein in the midrib and into the leaf's vascular network. Water then
traverses the leaf from cell-to-cell and evaporates into the air spaces. How does water travel from cell-to-cell? In plant tissues like
leaf mesophyll, neighbouring cells are connected by pores called plasmodesmata. These are tiny pores that traverse the cell
walls and cell membranes of neighbouring cells. They are membrane-lined and the cytoplasms of neighbouring cells are
continuous as the cytoplasm fills the plasmodesmatal channels. Thus, the cytoplasms of plant cells form a connected continuous
system called the symplast. Water can travel through the cytoplasm from cell-to-cell through the symplast. Water can, however,
also move through the extracellular spaces and cell walls that together form the apolast. The cell walls are principally made of
cellulose microfibrils which form a porous mesh. Water can move through the apoplast by capillarity (and probably moves faster
than through the symplast).
losing too much water. Stomata will open and close according to a circadian rhythm as well as in direct response to light and dark.
Stomata may also close in response to wounding - plants can lose a lot of water through open wounds and some plants, e.g.
tomato plants, react rapidly to damage by transmitting electrical signals throughout their leaves which trigger the stomata to close.
Despite these measures, plants lose a lot of water through their stomata and this water needs to be replaced. Xylem vessels carry
water from the roots, up the stem and to the leaves. Plants exploit this situation, making the most of a 'bad thing' and they utilise
the so-called transpiration stream of sap ascending in the xylem to carry mineral nutrients, obtained by the roots from the soil,
with it. Transpiration from the leaves also helps to keep them cool (important as they are purposefully exposed to the sunlight!).
The xylem sap is carried into each leaf, along the central vein in the midrib and into the leaf's vascular network. Water then
traverses the leaf from cell-to-cell and evaporates into the air spaces. How does water travel from cell-to-cell? In plant tissues like
leaf mesophyll, neighbouring cells are connected by pores called plasmodesmata. These are tiny pores that traverse the cell
walls and cell membranes of neighbouring cells. They are membrane-lined and the cytoplasms of neighbouring cells are
continuous as the cytoplasm fills the plasmodesmatal channels. Thus, the cytoplasms of plant cells form a connected continuous
system called the symplast. Water can travel through the cytoplasm from cell-to-cell through the symplast. Water can, however,
also move through the extracellular spaces and cell walls that together form the apolast. The cell walls are principally made of
cellulose microfibrils which form a porous mesh. Water can move through the apoplast by capillarity (and probably moves faster
than through the symplast).
Capillarity
Water will move upwards inside a very narrow glass tube (capillary tube) for a short distance. Water does this because it is
sticky. Water molecules are electrically charged and dipolar (one end or pole of the molecule is negatively charged, the other
pole positively charged). Opposite charges attract and water molecules bond or stick to each other (by hydrogen bonds in a
process called cohesion - like sticking to like) and they are also electrically attracted to other materials, like the glass wall of a
capillary tube, or the cellulose microfibrils of the apoplast (in a process called adhesion - two unlike materials sticking
together). This attraction pulls water along inside narrow spaces (this is how a sponge can passively soak-up water). The
force that drives this movement of water is the capillary force of cohesion-adhesion. This force drives the movement of water
through plant apoplasts. Capillarity is of key importance in water transport along the outside of moss stems in ectohydric
mosses.
Water moves from the xylem across the leaf to the air spaces by the apoplast and symplast and then evaporates through the
stomata (transpiration).
Water will move upwards inside a very narrow glass tube (capillary tube) for a short distance. Water does this because it is
sticky. Water molecules are electrically charged and dipolar (one end or pole of the molecule is negatively charged, the other
pole positively charged). Opposite charges attract and water molecules bond or stick to each other (by hydrogen bonds in a
process called cohesion - like sticking to like) and they are also electrically attracted to other materials, like the glass wall of a
capillary tube, or the cellulose microfibrils of the apoplast (in a process called adhesion - two unlike materials sticking
together). This attraction pulls water along inside narrow spaces (this is how a sponge can passively soak-up water). The
force that drives this movement of water is the capillary force of cohesion-adhesion. This force drives the movement of water
through plant apoplasts. Capillarity is of key importance in water transport along the outside of moss stems in ectohydric
mosses.
Water moves from the xylem across the leaf to the air spaces by the apoplast and symplast and then evaporates through the
stomata (transpiration).


Click here and here
for more information on the structure of xylem and wood.
It is the transpiration of water from leaves which is the main driving force for the movement of
water in xylem. Loss of water from the leaves creates a negative suction pressure that draws
water up the stem. Peak flow rates in xylem are about 1 mm/s, though maximum velocities as high
as 0.8 m/s have been reported. Flow in the xylem stops at night and then velocities rise in the
morning, peaking around midday. Wider vessels have larger maximum velocities but are more
prone to cavitation (the formation of air bubbles that block xylem vessels) in cold weather. For
this reason, the evergreen cold-tolerant conifers have narrower vessels and deciduous trees
produce narrower vessels in Autumn and wider vessels in Spring and Summer - this annual cycle
in vessel size creates the annual growth ring seen in trees. Cavitation occurs when the water
column breaks - the water is being pulled up to a great height and may break under its won
weight, especially if cold and 'brittle' and fracture of the water column is also more likely in wider
vessels. The fact that columns of water can be lifted up against gravity at all is due to cohesion:
water molecules stick to one another and so they move up the xylem as a continuous column.
Water will also adhere to the inside of the vessel wall. This physical explanation of xylem sap
ascent is called the cohesion-tension theory, as the water is pulled up the tree (and so is in
tension). The movement of sap in the xylem is called the transpiration stream.
It is the transpiration of water from leaves which is the main driving force for the movement of
water in xylem. Loss of water from the leaves creates a negative suction pressure that draws
water up the stem. Peak flow rates in xylem are about 1 mm/s, though maximum velocities as high
as 0.8 m/s have been reported. Flow in the xylem stops at night and then velocities rise in the
morning, peaking around midday. Wider vessels have larger maximum velocities but are more
prone to cavitation (the formation of air bubbles that block xylem vessels) in cold weather. For
this reason, the evergreen cold-tolerant conifers have narrower vessels and deciduous trees
produce narrower vessels in Autumn and wider vessels in Spring and Summer - this annual cycle
in vessel size creates the annual growth ring seen in trees. Cavitation occurs when the water
column breaks - the water is being pulled up to a great height and may break under its won
weight, especially if cold and 'brittle' and fracture of the water column is also more likely in wider
vessels. The fact that columns of water can be lifted up against gravity at all is due to cohesion:
water molecules stick to one another and so they move up the xylem as a continuous column.
Water will also adhere to the inside of the vessel wall. This physical explanation of xylem sap
ascent is called the cohesion-tension theory, as the water is pulled up the tree (and so is in
tension). The movement of sap in the xylem is called the transpiration stream.
shoot on
a tree or the stem or branch of an herbaceous plant)
the vascular tissue is arranged in bundles. The stem is covered
in a layer of cells forming an epidermis. Beneath this there are
often several layers of thick-walled cells called collenchyma
which strengthen the stem. Each bundle contains xylem on the
inside and another vascular tissue, the phloem on the outside.
Xylem vessels and phloem fibres also strengthen the stem.
the vascular tissue is arranged in bundles. The stem is covered
in a layer of cells forming an epidermis. Beneath this there are
often several layers of thick-walled cells called collenchyma
which strengthen the stem. Each bundle contains xylem on the
inside and another vascular tissue, the phloem on the outside.
Xylem vessels and phloem fibres also strengthen the stem.
Plant
Roots
Before considering the phloem transport system, let's look at water uptake in roots.
Before considering the phloem transport system, let's look at water uptake in roots.
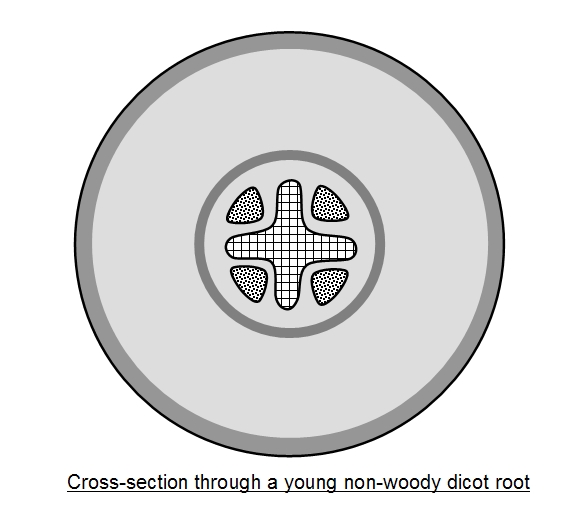
The
arrangement of the xylem and phloem in a
typical young (non-woody) dicotyledonous root is
shown on the left and below. Compare this pattern
with that found in the stem. The xylem is central, so
water must enter the root and travel across the
cortex and cross a cylinder of cells called the
endodermis in order to reach it.
typical young (non-woody) dicotyledonous root is
shown on the left and below. Compare this pattern
with that found in the stem. The xylem is central, so
water must enter the root and travel across the
cortex and cross a cylinder of cells called the
endodermis in order to reach it.
One
piece of experimental evidence for suction forces driving xylem
sap ascent in the stem comes from stem-girth
measurements. These show that at dawn the trunk contracts slightly, reaching its narrowest at midday and then
increases again to its night-time value. This corresponds to the stomata opening at dawn, as the tree begins
photosynthesis, creating a suction which draws the xylem vessels in, narrowing them slightly and this narrowing is
greatest when transpiration is greatest at around midday. More direct methods are available, such as heat-pulse
measurements. An electrical wire is placed just beneath the bark at one point of the stem and a thermocouple placed
about 4 cm further up. A brief electric current heats the wire and the xylem near it and then the time measured for
the heat pulse to travel up the stem can be measured, demonstrating the ascent of sap in the wood.
measurements. These show that at dawn the trunk contracts slightly, reaching its narrowest at midday and then
increases again to its night-time value. This corresponds to the stomata opening at dawn, as the tree begins
photosynthesis, creating a suction which draws the xylem vessels in, narrowing them slightly and this narrowing is
greatest when transpiration is greatest at around midday. More direct methods are available, such as heat-pulse
measurements. An electrical wire is placed just beneath the bark at one point of the stem and a thermocouple placed
about 4 cm further up. A brief electric current heats the wire and the xylem near it and then the time measured for
the heat pulse to travel up the stem can be measured, demonstrating the ascent of sap in the wood.
The
detailed structure of a sector of such a root is shown below:


In woody
stems, a ring of secondary xylem
or wood is added each year. The outer
rings of wood are the newest and only
these contain functional vessels. Older
vessels become blocked and
non-functional. The oldest parts form the
heartwood. Also within the wood are radial
plates of parenchyma cells called rays,
which can transport materials across the
stem, for example depositing waste
compounds into the heartwood giving it a
different colour. The bark has two main
layers - the inner fibrous layer of phloem
and the outer layer of cork cells.
or wood is added each year. The outer
rings of wood are the newest and only
these contain functional vessels. Older
vessels become blocked and
non-functional. The oldest parts form the
heartwood. Also within the wood are radial
plates of parenchyma cells called rays,
which can transport materials across the
stem, for example depositing waste
compounds into the heartwood giving it a
different colour. The bark has two main
layers - the inner fibrous layer of phloem
and the outer layer of cork cells.
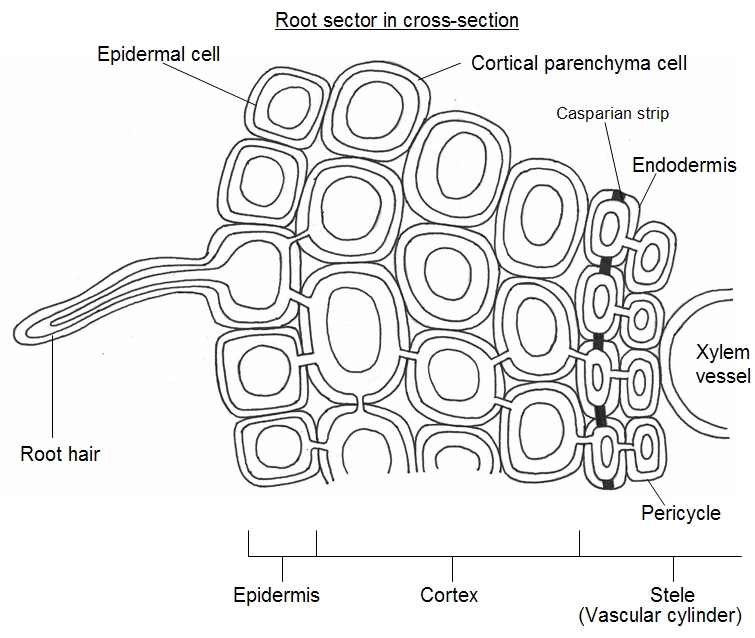
Some of
the root epidermal cells, situated near the growing root tips,
have projections called root hairs
(unicellular hairs) which increase the surface area of the root for water absorption, though other epidermal cells
may also absorb water. The diagram below shows two possible routes of this water absorption. The bottom arrow
(blue) shows water moving through plasmodesmata in the symplast and is the symplastic pathway. Note that to
reach the symplast the water first had to cross the cell membrane of the root hair cell. (See plant cell structure).
The top arrow (red) shows water being absorbed through the apoplast (apoplastic pathway) traveling through
the sponge-like meshes of the cell walls by capillarity, until it reaches a cylinder of cells called the endodermis.
(The apoplast pathway is probably faster than the symplast pathway). These cells have a hoop of water-proof
material in their cell walls (suberin) forming the Casparian strip. At this point the apolastic pathway is blocked
and the water is forced to detour along the dashed part of the arrow, which leads into the symplast. At this point
the water was forced to cross the cell membrane of the endodermis cell. This is the key point - whichever route
the water takes to reach the xylem, it has to cross at least one cell-surface membrane.
(unicellular hairs) which increase the surface area of the root for water absorption, though other epidermal cells
may also absorb water. The diagram below shows two possible routes of this water absorption. The bottom arrow
(blue) shows water moving through plasmodesmata in the symplast and is the symplastic pathway. Note that to
reach the symplast the water first had to cross the cell membrane of the root hair cell. (See plant cell structure).
The top arrow (red) shows water being absorbed through the apoplast (apoplastic pathway) traveling through
the sponge-like meshes of the cell walls by capillarity, until it reaches a cylinder of cells called the endodermis.
(The apoplast pathway is probably faster than the symplast pathway). These cells have a hoop of water-proof
material in their cell walls (suberin) forming the Casparian strip. At this point the apolastic pathway is blocked
and the water is forced to detour along the dashed part of the arrow, which leads into the symplast. At this point
the water was forced to cross the cell membrane of the endodermis cell. This is the key point - whichever route
the water takes to reach the xylem, it has to cross at least one cell-surface membrane.
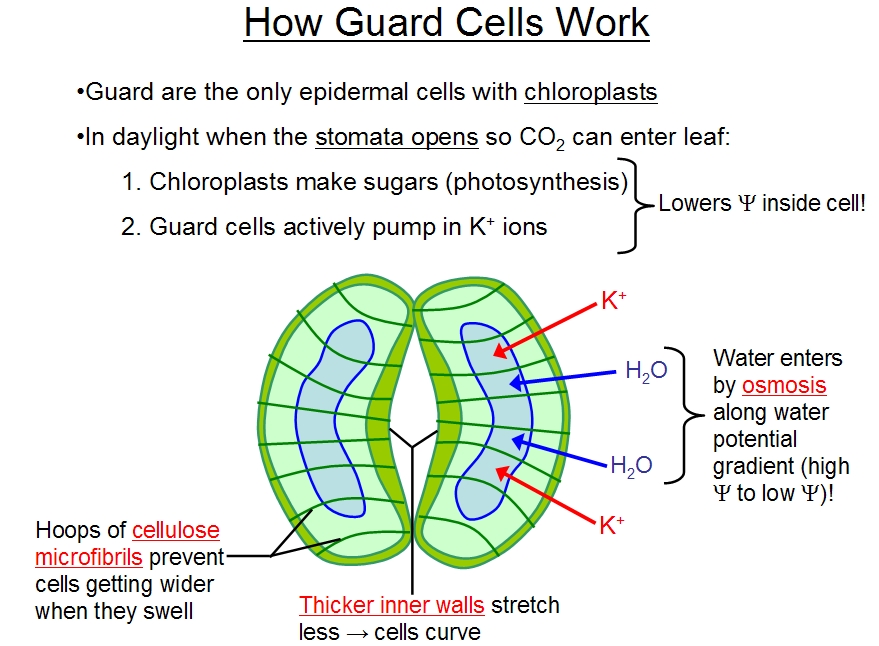
These
cell-surface membranes are able to control water entry and so the
root can control water and mineral
uptake at these points. Protein pumps in these membranes actively pump mineral salts, obtained from the soil
water, into their cytoplasm, using cellular energy in the form of ATP (adenosine trisphosphate). This loading with
salts creates an osmotic gradient (the water potential inside the cell is lowered - see below for an explanation of
water potential) which then drives water into the cell, across the membrane (as water moves from a region of low
solute gradient, or high water potential, to a region of high solute concentration, or low water potential). Thus, by
using energy to accumulate the dilute and much needed mineral slats from the soil, the cells are essentially
pumping water into the root and into the xylem! The xylem will then carry this water and these minerals (the
minerals are essential nutrients for the plant) in the transpiration stream. This pumping creates a positive
pressure in the root xylem, helping to push water up the stem - the so-called root pressure. This root pressure
may force water up by a metre or so and may be sufficient tod rive water transport in small plants, however, in tall
trees transpiration is by far the main driving force for xylem sap movement.
uptake at these points. Protein pumps in these membranes actively pump mineral salts, obtained from the soil
water, into their cytoplasm, using cellular energy in the form of ATP (adenosine trisphosphate). This loading with
salts creates an osmotic gradient (the water potential inside the cell is lowered - see below for an explanation of
water potential) which then drives water into the cell, across the membrane (as water moves from a region of low
solute gradient, or high water potential, to a region of high solute concentration, or low water potential). Thus, by
using energy to accumulate the dilute and much needed mineral slats from the soil, the cells are essentially
pumping water into the root and into the xylem! The xylem will then carry this water and these minerals (the
minerals are essential nutrients for the plant) in the transpiration stream. This pumping creates a positive
pressure in the root xylem, helping to push water up the stem - the so-called root pressure. This root pressure
may force water up by a metre or so and may be sufficient tod rive water transport in small plants, however, in tall
trees transpiration is by far the main driving force for xylem sap movement.
Water
potential (given the Greek letter psi as its symbol) is simply the
potential energy possessed by a unit
volume (a set volume, e.g. 1 metre cubed) of water. It is the sum total of the various kinds of potential energy
the water can possess. The most familiar is the gravitational potential energy. Take a ball and raise it in the air
and you have increased its gravitational potential energy. Energy is the ability to cause change and this
gravitational energy is potential energy because it has the potential to cause change, but will not do so until the
ball is released. When released, the ball falls to the ground as it loses gravitational potential energy which is
turned into kinetic energy (the energy of movement).
Water potential is a useful concept for describing water movement in plants. It is often said that water will
always move from a region of high water potential to a region of low water potential (down a water potential
gradient) - like the falling ball. However, this is not strictly true, water will move down a water potential gradient
of its accord if allowed to do so. However, it is possible to move water from a region of low water potential to a
region of high water potential, by supplying energy (as you did when raising the ball) - the water will then gain
potential energy. In xylem water does move from high to low water potential. The plant expends none of its own
energy to move the water, it simply opens the stomata and transpiration does the rest and water moves from
high water potential in the roots to low water potential in the air above the canopy. (transpiration creates a
negative pressure potential at the top of the tree, sucking the water up the stem). However, in phloem water
can move from low to high water potential because the plant uses cellular energy (in the form of ATP) to
actively pump water up or down the stem (it creates positive pressure that pushes the water along in the
phloem). In both xylem and phloem it is the pressure which drives water movement.
The phloem contains a system of vessels for transporting photoassimilates (the products of photosynthesis
like amino acids and sugars) around the plant. Sugar is transported mainly as sucrose and nitrogen as amino
acids (the building blocks of proteins). The reason why sucrose is transported rather than glucose is probably
because sucrose is harder for bacteria to metabolise and so transporting sucrose reduces the risk of infection.
(Similarly insects transport the sugar trehalose and seaweeds mannitol. Mammals, however, transport glucose
in their blood stream since this can be rapidly utilised by nerve cells and having such large brains necessitates
a ready fuel supply. This, combined with their constantly warm temperatures, makes mammals very prone to
infection and so they have evolved highly sophisticated immune systems. Still, however, major wounds to large
animals in nature often result in death, not from the wound, but from the ensuing infection).
Phloem is a tissue comprising several different cell types, including parenchyma, tough sclerenchyma fibres,
and the phloem vessels, called sieve tubes, which are made of sieve-tube members or elements (sieve tube
cells). Phloem carries sugars (and other photoassimilates) from sources where they are released (e.g.
photosynthesising leaves where they are made, or storage organs like bulbs germinating in Spring) to sinks
where they used (e.g. growing fruit, shaded leaves, storage organs like roots which store the sugars as starch).
This process is called translocation (a name sometimes also used to describe movement of materials in xylem).
It is now known that it is not just the sugars and other solutes that move through the phloem, but the water
moves with it - the whole phloem sap moves in bulk flow (mass flow or mass transport).
In photosynthesising leaves, the sugars can be transported toward the phloem sieve tubes in the veins by
either the symplast or apoplast pathways, as shown below:
volume (a set volume, e.g. 1 metre cubed) of water. It is the sum total of the various kinds of potential energy
the water can possess. The most familiar is the gravitational potential energy. Take a ball and raise it in the air
and you have increased its gravitational potential energy. Energy is the ability to cause change and this
gravitational energy is potential energy because it has the potential to cause change, but will not do so until the
ball is released. When released, the ball falls to the ground as it loses gravitational potential energy which is
turned into kinetic energy (the energy of movement).
Water potential is a useful concept for describing water movement in plants. It is often said that water will
always move from a region of high water potential to a region of low water potential (down a water potential
gradient) - like the falling ball. However, this is not strictly true, water will move down a water potential gradient
of its accord if allowed to do so. However, it is possible to move water from a region of low water potential to a
region of high water potential, by supplying energy (as you did when raising the ball) - the water will then gain
potential energy. In xylem water does move from high to low water potential. The plant expends none of its own
energy to move the water, it simply opens the stomata and transpiration does the rest and water moves from
high water potential in the roots to low water potential in the air above the canopy. (transpiration creates a
negative pressure potential at the top of the tree, sucking the water up the stem). However, in phloem water
can move from low to high water potential because the plant uses cellular energy (in the form of ATP) to
actively pump water up or down the stem (it creates positive pressure that pushes the water along in the
phloem). In both xylem and phloem it is the pressure which drives water movement.
The phloem contains a system of vessels for transporting photoassimilates (the products of photosynthesis
like amino acids and sugars) around the plant. Sugar is transported mainly as sucrose and nitrogen as amino
acids (the building blocks of proteins). The reason why sucrose is transported rather than glucose is probably
because sucrose is harder for bacteria to metabolise and so transporting sucrose reduces the risk of infection.
(Similarly insects transport the sugar trehalose and seaweeds mannitol. Mammals, however, transport glucose
in their blood stream since this can be rapidly utilised by nerve cells and having such large brains necessitates
a ready fuel supply. This, combined with their constantly warm temperatures, makes mammals very prone to
infection and so they have evolved highly sophisticated immune systems. Still, however, major wounds to large
animals in nature often result in death, not from the wound, but from the ensuing infection).
Phloem is a tissue comprising several different cell types, including parenchyma, tough sclerenchyma fibres,
and the phloem vessels, called sieve tubes, which are made of sieve-tube members or elements (sieve tube
cells). Phloem carries sugars (and other photoassimilates) from sources where they are released (e.g.
photosynthesising leaves where they are made, or storage organs like bulbs germinating in Spring) to sinks
where they used (e.g. growing fruit, shaded leaves, storage organs like roots which store the sugars as starch).
This process is called translocation (a name sometimes also used to describe movement of materials in xylem).
It is now known that it is not just the sugars and other solutes that move through the phloem, but the water
moves with it - the whole phloem sap moves in bulk flow (mass flow or mass transport).
In photosynthesising leaves, the sugars can be transported toward the phloem sieve tubes in the veins by
either the symplast or apoplast pathways, as shown below:
Note
that the apoplastic pathway involves the sugars crossing the cell
membranes of specialised parenchyma
cells called companion cells (there is typically one companion cell per sieve element). This cell-surface
membrane can regulate the transport of the sugar and also drives it by actively pumping sugar from the
neighbouring parenchyma cells into the phloem sieve tubes (using membrane protein pumps that require
cellular energy in the form of ATP). This pumping of sugars into the companion cell/sieve tube element
creates an osmotic gradient, or put another way, lowers the solute potential of the companion cell and hence
lowers its water potential. (Solute potential is highest for pure water which is given the value zero, so other
solutions will have a lower or more negative solute potential). Water then follows the sugar as it moves from
high to low water potentials and enters the sieve tubes. As the phloem travels along the sieve tubes in the
source, passing from element to element, it is given a push as more sugars are loaded in, creating a pulsatile
pressure (positive pressure potential) that pushes the phloem along.
Unloading of the sugars occurs at the sink. Here companion cells actively pump the sugars out from the
phloem, causing water to follow. This unloading adds to the pressure gradient in the sieve tubes - the phloem
is pushed along by sugar loading at the source and pulled along by sugar unloading at the sink. This
movement from source to sink can occur in any direction within the plant, but one of the major routes will be
from the leaves, down the stem, to the roots. Roots can not and do not photosynthesise and need sugars to
meet their own energy needs, they also store excess sugars as starch, safely underground and away from
browsing herbivores!
cells called companion cells (there is typically one companion cell per sieve element). This cell-surface
membrane can regulate the transport of the sugar and also drives it by actively pumping sugar from the
neighbouring parenchyma cells into the phloem sieve tubes (using membrane protein pumps that require
cellular energy in the form of ATP). This pumping of sugars into the companion cell/sieve tube element
creates an osmotic gradient, or put another way, lowers the solute potential of the companion cell and hence
lowers its water potential. (Solute potential is highest for pure water which is given the value zero, so other
solutions will have a lower or more negative solute potential). Water then follows the sugar as it moves from
high to low water potentials and enters the sieve tubes. As the phloem travels along the sieve tubes in the
source, passing from element to element, it is given a push as more sugars are loaded in, creating a pulsatile
pressure (positive pressure potential) that pushes the phloem along.
Unloading of the sugars occurs at the sink. Here companion cells actively pump the sugars out from the
phloem, causing water to follow. This unloading adds to the pressure gradient in the sieve tubes - the phloem
is pushed along by sugar loading at the source and pulled along by sugar unloading at the sink. This
movement from source to sink can occur in any direction within the plant, but one of the major routes will be
from the leaves, down the stem, to the roots. Roots can not and do not photosynthesise and need sugars to
meet their own energy needs, they also store excess sugars as starch, safely underground and away from
browsing herbivores!



Above:
The structure of a phloem sieve tube and its companion
cells. Note the peripheral layer of cytoplasm against the walls of
the sieve tube members (sieve tube elements, sieve elements
or sieve tube cells) bounded by a degraded tonoplast (the
membrane around the original cell vacuole) which loosely
separates the cytoplasm from the lumen. During differentiation,
condensed and confined to the margins of the tube and may be
condensed into P-protein bodies. Smooth endoplasmic
reticulum (SER) gradually degenerates and may be absent
altogether in mature sieve tube elements.
cells. Note the peripheral layer of cytoplasm against the walls of
the sieve tube members (sieve tube elements, sieve elements
or sieve tube cells) bounded by a degraded tonoplast (the
membrane around the original cell vacuole) which loosely
separates the cytoplasm from the lumen. During differentiation,
condensed and confined to the margins of the tube and may be
condensed into P-protein bodies. Smooth endoplasmic
reticulum (SER) gradually degenerates and may be absent
altogether in mature sieve tube elements.
Adjacent
sieve tube members are separated by porous sieve plates with pore
diameters ranging from about 1 micrometre to 14 micrometres.
responsible for providing most of the energy needed to pump materials into and out of the phloem tubes to neighbouring parenchyma cells.
Large single plasmodesmata and groups of plasmodesmata, called sieve areas, connect the cytoplasm of the companion cell to its sieve tube
element. 'Sieve area' is a general term for an area of pores connecting a sieve tube cell to another cell. When these pores are especially
large and confined to a distinct piece of cell wall, as in the end-walls of the sieve tube cells, they are called sieve plates.
When a sieve tube is cut or wounded, the P-protein spreads out (unwinds) rapidly to fill the lumen and block the pores in the sieve plates, so
blocking the vessel and preventing further loss of phloem sap.
responsible for providing most of the energy needed to pump materials into and out of the phloem tubes to neighbouring parenchyma cells.
Large single plasmodesmata and groups of plasmodesmata, called sieve areas, connect the cytoplasm of the companion cell to its sieve tube
element. 'Sieve area' is a general term for an area of pores connecting a sieve tube cell to another cell. When these pores are especially
large and confined to a distinct piece of cell wall, as in the end-walls of the sieve tube cells, they are called sieve plates.
When a sieve tube is cut or wounded, the P-protein spreads out (unwinds) rapidly to fill the lumen and block the pores in the sieve plates, so
blocking the vessel and preventing further loss of phloem sap.
Gymnosperms,
including conifers like the pine tree, do not have sieve tubes.
Instead they have sieve cells that are connected together by
sieve areas with smaller pores (the pores are generally less than 0.8 micrometres in diameter), and so do not form an open tube. Sieve cells
are much narrower (less than about 5 micrometres in diameter, compared to sieve tube cells which are up to about 50 micrometres in
diameter) and several times longer than sieve tube cells. Specialised parenchyma cells function much like companion cells, but are called
albuminous cells in gymnosperms. (In development, a sieve tube cell and its companion cell are produced from the same parent cell by cell
division, whereas sieve cells and albuminous cells do not have a common parent cell). Sieve cells in gymnosperms retain their SER (smooth
endoplasmic reticulum) as a network that terminates on either side of the sieve are pores, and do not have P-protein. When a gymnosperm
sieve cell is wounded, the SER swells up and expands to block the pores. This SER network may also have some role in transport and it is
possible that translocation in gymnosperm phloem operates by a different mechanism to that described for angiosperm sieve tubes.
Mosses and algae also have phloem-like tissues. In mosses elongated leptoid cells are connected by fields of plasmodesmata (with pore
diameters of 0.12 to 0.15 micrometres in Dendroligotricum) and so resemble sieve cells. In seaweeds, trumpet hyphae are made up of sieve
cells connected by sieve areas or sieve plates, with pore diameters ranging from 0.04-0.09 micrometres in Laminaria, to 6.0 micrometres in
the giant kelp Macrocystis. In gymnosperms the pore diameter is typically less than 0.8 micrometres, and 6.5 micrometres in Fagus (beech).
The general pattern seems to be that as plants moved onto land and grew larger, so did their pore sizes, which reduces resistance to sap
movement and increases the maximum rate of translocation in a vessel of a given diameter. Gymnosperms may form very large trees and are
at odds with this trend and more will be said about this later.
sieve areas with smaller pores (the pores are generally less than 0.8 micrometres in diameter), and so do not form an open tube. Sieve cells
are much narrower (less than about 5 micrometres in diameter, compared to sieve tube cells which are up to about 50 micrometres in
diameter) and several times longer than sieve tube cells. Specialised parenchyma cells function much like companion cells, but are called
albuminous cells in gymnosperms. (In development, a sieve tube cell and its companion cell are produced from the same parent cell by cell
division, whereas sieve cells and albuminous cells do not have a common parent cell). Sieve cells in gymnosperms retain their SER (smooth
endoplasmic reticulum) as a network that terminates on either side of the sieve are pores, and do not have P-protein. When a gymnosperm
sieve cell is wounded, the SER swells up and expands to block the pores. This SER network may also have some role in transport and it is
possible that translocation in gymnosperm phloem operates by a different mechanism to that described for angiosperm sieve tubes.
Mosses and algae also have phloem-like tissues. In mosses elongated leptoid cells are connected by fields of plasmodesmata (with pore
diameters of 0.12 to 0.15 micrometres in Dendroligotricum) and so resemble sieve cells. In seaweeds, trumpet hyphae are made up of sieve
cells connected by sieve areas or sieve plates, with pore diameters ranging from 0.04-0.09 micrometres in Laminaria, to 6.0 micrometres in
the giant kelp Macrocystis. In gymnosperms the pore diameter is typically less than 0.8 micrometres, and 6.5 micrometres in Fagus (beech).
The general pattern seems to be that as plants moved onto land and grew larger, so did their pore sizes, which reduces resistance to sap
movement and increases the maximum rate of translocation in a vessel of a given diameter. Gymnosperms may form very large trees and are
at odds with this trend and more will be said about this later.


which
expand to fill all the space right up to the rigid cell walls
('boxes') that contain them. This will happen if the cells are
exposed to a
sugar solution of low concentration - water will move into the cells until the cells are full. Right - cells which have lost water by osmosis,
causing their protoplasts to shrink away from the cell walls, a phenomenon called plasmolysis (the cells are said to be plasmolysed). This
happened because the tissue was soaked in a sugar solution of high concentration, causing water to diffuse out from the cells by osmosis.
As water makes up some 70% of the volume of cells, the protoplasts have shrunk and come away from their surrounding cell walls.
This phenomenon is important in plants. In soft green or fleshy plant parts, when the plant has sufficient water, the cells are full and the
protoplasts push against the cell walls, keeping them rigid (rather like tyres filled with water at high pressure) and giving the plant parts
support - the cells and plant parts are said to be turgid (swollen and rigid). This is important for keeping leaves and green stems upright to
intercept sunlight - the tissues (especially parenchyma) functions as a pressurised cellular solid. A common example of a pressurised
cellular solid is polystyrene - tiny balls of foam filled with air which when packed together become rigid - bending the polystyrene slightly
squeezes the air inside the balls which resist bending. If a plant is dehydrated (due to water loss by evaporation) however, the cells
plasmolyse and lose pressure and the cell walls lose rigidity and the cells become flaccid and the plant wilts. This is also protective - as
leaves droop so they become less-exposed to the drying sun and/or wind. Of course, the plant cannot survive in this desiccated state
indefinitely, as water is vital to cell chemistry, but they can recover quickly if the plant receives water.
Animal cells, such as mammalian red blood cells, similarly shrink if they lose water by osmosis to a concentrated solution. However, if
immersed in distilled water, which is fairly pure, then water enters the cells by osmosis, causing them to swell and burst - mammalian cells
will explode within seconds on contact with distilled water. Single-celled organisms that resemble animal cells, like amoeba, can survive
because they have contractile vacuoles, which swell up with water, as excess water enters the cell, and then contract to expel the water.
sugar solution of low concentration - water will move into the cells until the cells are full. Right - cells which have lost water by osmosis,
causing their protoplasts to shrink away from the cell walls, a phenomenon called plasmolysis (the cells are said to be plasmolysed). This
happened because the tissue was soaked in a sugar solution of high concentration, causing water to diffuse out from the cells by osmosis.
As water makes up some 70% of the volume of cells, the protoplasts have shrunk and come away from their surrounding cell walls.
This phenomenon is important in plants. In soft green or fleshy plant parts, when the plant has sufficient water, the cells are full and the
protoplasts push against the cell walls, keeping them rigid (rather like tyres filled with water at high pressure) and giving the plant parts
support - the cells and plant parts are said to be turgid (swollen and rigid). This is important for keeping leaves and green stems upright to
intercept sunlight - the tissues (especially parenchyma) functions as a pressurised cellular solid. A common example of a pressurised
cellular solid is polystyrene - tiny balls of foam filled with air which when packed together become rigid - bending the polystyrene slightly
squeezes the air inside the balls which resist bending. If a plant is dehydrated (due to water loss by evaporation) however, the cells
plasmolyse and lose pressure and the cell walls lose rigidity and the cells become flaccid and the plant wilts. This is also protective - as
leaves droop so they become less-exposed to the drying sun and/or wind. Of course, the plant cannot survive in this desiccated state
indefinitely, as water is vital to cell chemistry, but they can recover quickly if the plant receives water.
Animal cells, such as mammalian red blood cells, similarly shrink if they lose water by osmosis to a concentrated solution. However, if
immersed in distilled water, which is fairly pure, then water enters the cells by osmosis, causing them to swell and burst - mammalian cells
will explode within seconds on contact with distilled water. Single-celled organisms that resemble animal cells, like amoeba, can survive
because they have contractile vacuoles, which swell up with water, as excess water enters the cell, and then contract to expel the water.
Comment on this article!
Osmosis
and Plasmolysis
Not all scientists define water potential in the same way. We have adopted the definition in terms of potential energy per unit volume. More or
less equivalently, some define it as potential energy per mole (one mole being about 6.022 x 10^23 particles, or molecules of water in this
case). Other definitions refer to water potential as the free energy of water per unit volume or per mole. Free energy is the energy available
to do work, and so is essentially equivalent to potential energy in most cases. However, free energy is sometimes defined as internal energy
only. A system, such as a mass of water, has both internal energy (due to the movements of its molecules relative to each other) and
external energy (due to the movement of the body of water as a whole). In plant biology, we need to include both external and internal
energy - external energy is important when we consider movement of water up a tree - as it ascends, the water as a whole gains gravitational
potential energy, which has the potential to do work should the water be released and allowed to fall to the ground. Internal energy is
important as this drives diffusion.
Diffusion is the net or overall movement of molecules from a region of higher concentration to a region of lower concentration. (By
concentration we mean the number of molecules in a given volume, such as the number of molecules per cubic meter). This movement
occurs because molecules are in constant thermal motion (they move about and the hotter they are, the more they move) and this
movement is random. If a membrane separates the regions of higher and lower concentration, then diffusion will only take place if the
membrane is permeable to the substance diffusing - for example if it is porous and the pores are large enough to allow the molecules
through. The cell-surface membrane works in this way - it is a selectively-permeable membrane, allowing only certain substances to cross it.
Water is one such substance. However, because cell membranes are made principally of lipids (fats/oils) in the form of phospholipids, water
does not cross easily (oil and water do not mix - water does not dissolve well in fat and vice versa) and will only slowly leak across a
phospholipid membrane. However, cells need water and so cell membranes have protein pores, called aquaporins, that are the right size to
allow water molecules to cross easily. This is an example of facilitated diffusion - the protein pores help or facilitate the diffusion of water
across the membrane. Diffusion of water across cell membranes is so important that it is given its own special name - osmosis.
In biology we rarely deal with 'pure' water, since biological fluids are all solution. A solution is a mixture of materials, in which certain
substances, called solutes, are dissolved in a solvent (which is the main component of the mixture). Most often in biology the solvent of
interest is water and this will contain a variety of dissolved solutes such as salts (like sodium chloride or common table salt, NaCl, and slats of
potassium, calcium and iron and other metals) sugars and amino acids. When salt, NaCl, dissolves in water, it splits up into ions of sodium,
Na+ and chloride, Cl- (an ion is an atom or molecule that gains one or more units of net electric charge, either positive or negative). In such
a solution it is inconvenient to think of the concentration of water as this is hard to measure, and we usually think in terms of concentration of
solute. Since ions, atoms or molecules of solvent occupy space that would otherwise be occupied by water molecules, the higher the total
solute concentration (of all solutes) the lower the concentration of water. Thus water will diffuse from a region of lower solute concentration
(higher water concentration) to a region of higher solute concentration (lower water concentration).
Osmosis across plant cell membranes can be easily demonstrated using plant tissues. A classic experiment involves taking strips of onion
epidermis (such as the translucent tissue-paper like layer on the inside of the fleshy leaves of the onion bulb which is one-cell thick and so
easily observed under the light microscope) and immersing them for twenty minutes or so in different concentrations of solution, typically a
solution of sucrose sugar. Some results from such an experiment are shown below:
Not all scientists define water potential in the same way. We have adopted the definition in terms of potential energy per unit volume. More or
less equivalently, some define it as potential energy per mole (one mole being about 6.022 x 10^23 particles, or molecules of water in this
case). Other definitions refer to water potential as the free energy of water per unit volume or per mole. Free energy is the energy available
to do work, and so is essentially equivalent to potential energy in most cases. However, free energy is sometimes defined as internal energy
only. A system, such as a mass of water, has both internal energy (due to the movements of its molecules relative to each other) and
external energy (due to the movement of the body of water as a whole). In plant biology, we need to include both external and internal
energy - external energy is important when we consider movement of water up a tree - as it ascends, the water as a whole gains gravitational
potential energy, which has the potential to do work should the water be released and allowed to fall to the ground. Internal energy is
important as this drives diffusion.
Diffusion is the net or overall movement of molecules from a region of higher concentration to a region of lower concentration. (By
concentration we mean the number of molecules in a given volume, such as the number of molecules per cubic meter). This movement
occurs because molecules are in constant thermal motion (they move about and the hotter they are, the more they move) and this
movement is random. If a membrane separates the regions of higher and lower concentration, then diffusion will only take place if the
membrane is permeable to the substance diffusing - for example if it is porous and the pores are large enough to allow the molecules
through. The cell-surface membrane works in this way - it is a selectively-permeable membrane, allowing only certain substances to cross it.
Water is one such substance. However, because cell membranes are made principally of lipids (fats/oils) in the form of phospholipids, water
does not cross easily (oil and water do not mix - water does not dissolve well in fat and vice versa) and will only slowly leak across a
phospholipid membrane. However, cells need water and so cell membranes have protein pores, called aquaporins, that are the right size to
allow water molecules to cross easily. This is an example of facilitated diffusion - the protein pores help or facilitate the diffusion of water
across the membrane. Diffusion of water across cell membranes is so important that it is given its own special name - osmosis.
In biology we rarely deal with 'pure' water, since biological fluids are all solution. A solution is a mixture of materials, in which certain
substances, called solutes, are dissolved in a solvent (which is the main component of the mixture). Most often in biology the solvent of
interest is water and this will contain a variety of dissolved solutes such as salts (like sodium chloride or common table salt, NaCl, and slats of
potassium, calcium and iron and other metals) sugars and amino acids. When salt, NaCl, dissolves in water, it splits up into ions of sodium,
Na+ and chloride, Cl- (an ion is an atom or molecule that gains one or more units of net electric charge, either positive or negative). In such
a solution it is inconvenient to think of the concentration of water as this is hard to measure, and we usually think in terms of concentration of
solute. Since ions, atoms or molecules of solvent occupy space that would otherwise be occupied by water molecules, the higher the total
solute concentration (of all solutes) the lower the concentration of water. Thus water will diffuse from a region of lower solute concentration
(higher water concentration) to a region of higher solute concentration (lower water concentration).
Osmosis across plant cell membranes can be easily demonstrated using plant tissues. A classic experiment involves taking strips of onion
epidermis (such as the translucent tissue-paper like layer on the inside of the fleshy leaves of the onion bulb which is one-cell thick and so
easily observed under the light microscope) and immersing them for twenty minutes or so in different concentrations of solution, typically a
solution of sucrose sugar. Some results from such an experiment are shown below:
Other related pages on Cronodon: Wood Plant Stems Leaves Plant Vascular Architecture |