| |
|
|
|
|
|
|
|
| Jungermanniales
- the leafy liverworts |
order of liverworts and contrast with thalloid liverworts such as Marchantia and Pellia and with
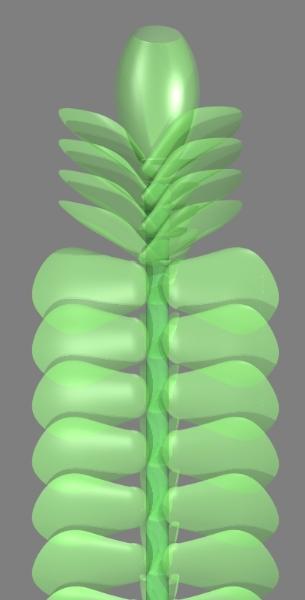
mosses and hornworts which collectively make up the bryophytes. The model above shows a
female shoot of a typical leafy liverwort. These plants, at most a few centimetres long, are
frequently found as epiphytes (growing on other plants).
Bryophytes are considered as 'primitive' plants, meaning that anatomically they are considered
more ancient forms, their common ancestor having evolved before that of flowering plants. They
show few anatomical adaptations to life on land, but have evolved the curious ability to survive
desiccation: they can apparently dry up completely, appearing dead, only to spring back to life
once water returns. Thus, they are highly adapted and highly evolved in their own ways. They are
generally tiny to small organisms, the shoots of leafy liverworts are typically 0.5 to 2 mm in
diameter, including the leaves.
The leaves are not the same as those of higher plants, lacking the intricate networks of veins and
the various layers of epithelial, mesophyll and vascular tissue. In leafy liverworts they rarely have
midribs and usually consist of a single layered sheet of photosynthetic cells. In those few forms in
which a midrib (vitta) is present, it resembles that in many mosses, consisting of elongated cells
which serve to accelerate the transport of foods made by photosnythesis (photoassimilates). Thus,
the leaves are not true leaves in the sense of flowering plants, and I am tempted to call them
'bryophylls'. There are no obvious supporting cells in the leaf, except that many of the cells have
thickened corner walls and when elongated midrib cells are present then these may also aid
support. otherwise, the turgor pressure of the cells is the principle support, the leaf tissue acting as
a 'pressurised cellular solid'.




liverworts. The stem generally has three rows of leaves along its length. However, in most,
especially those which have prostrate stems (stems lying down along the substrate) the leaves of
one of these rows is much reduced in size and often have a distinctly different shape and are
called amphigastria. These form the ventral row (underneath the stem) and associated with each
amphigastrum there maybe a few anchoring rhizoids (seen above when the model is tilted). The
two remaining rows occur on the sides (lateral rows). The rhizoids are generally extensions of
single-cells in liverworts and serve primarily for anchorage (very little transport of water occurs
within them and at a very slow rate). Thus the rhizoids are not rue roots. The rhizoids are usually
whitish turning browner with age, or sometimes purple.
Shoots may be vegetative, ending in a cluster of expanding leaves which protect the growing tip or
they may be fertile shoots, ending in female or male reproductive organs. The female shoot shown
here ends in a vase-like perianth which encloses and protects the female reproductive organs
(archegonia). The perianth consists of two or three modified leaves which may or may not be
fused together along their margins and is found in most leafy liverworts. The perianth is usually
surrounded at its base by modified leaves, called bracts, and modified amphigastria, called
bracteoles. The bracts may be larger or smaller than normal vegetative leaves. Branching of the
stem is common and may follow a wide variety of different patterns or may be apparently random.
Male side-branches are often short, compressed and catkin-like, with the modified scale-like leaves
sheltering the male reproductive organs (antheridia)
dorsal (topside) view. Note that in this prostrate stem, the leaves are tilted to maximise the
interception of light for photosynthesise. During growth the leaf emerges at right angles to the
stem (as if the shoot was erect) and then growth of stem tissue twists the leaf around into its final
position. This triple arrangement of leaves arises because the shoot grows from a single apical
meristem cell at its tip which acts as a meristem: this cell divides (by mitosis) and its daughters
give rise to the various tissues and organs. Since this cell is tetrahedral, like a triangular-based
pyramid, but upside-down it has three faces from which daughter cells separate, each in turn. This
gives rise to the three rows of leaves, with the row underneath those forms with prostrate stems
being greatly reduced to amphigastria.

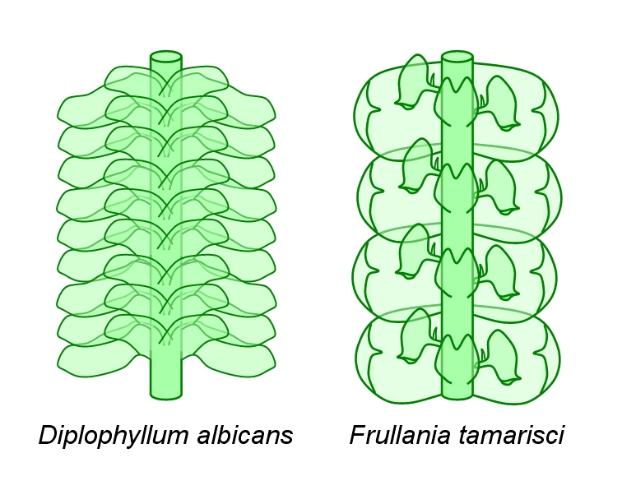
species. Above: left, when the rear edge of each leaf overlaps the front edge of the leaf behind it,
then we have the succubous arrangement (as in Lophocolea, for example). Right, when the front
edge of each leaf overlaps the back edge of the leaf in front of it, then we have the incubous
arrangement (as in Bazzania, for example).
leaves have two-pointed tips (sometimes three or four, sometimes only one rounded tip). In some
species, these two tips grow separately for much of the leaf's length, resulting in a bilobed leaf.
For example, in Diplophyllum ('double leaf', above left: dorsal view) and the basal part of the leaf
becomes folded with the smaller antical lobe lying across the larger postical lobe.
In Frullania (above right: ventral view) the smaller postical lobe forms a small helmet-shaped
pitcher, attached to a short stalk equipped with a spur. It is thought that these pitchers serve to
store water. In the tropical Colura and Pleurozia, the pitchers are effective at trapping minute
animals and protozoa and these plants may be carnivorous. It could also be that the pitchers
house symbionts, bacteria or protozoa (which may feed on the bacteria) which are beneficial to the
plant in some way. In the tropical Polyotus, both the lateral leaves and amphigastria form pitchers.
In some species, the leaves divide into filaments, which may resemble filamentous algae.
gametophyte, the haploid form of the
leafy liverworts which produces gametes.
(Haploid means that it ha sonly one set of
chromosomes rather than the usual two).
On the left is a female shoot tip of
Lophozia ventricosa. The female organs
are called archegonia. Each
archegonium arises from a single
surface cell and is a minute flask-shaped
structure, with a rounded base or venter
containing the single egg cell and a
ventral canal cell on top of the egg,
and then a narrow neck whose canal is
initially plugged by 5 (in most
Jungermanniales) neck canal cells. The
whole may be borne on a short stalk. A
group of these archegonia will typically be
produced at the tip of a female shoot,
encloded by the large parianth.
The male organs, the antheridia develop
on the male branches. Each antheridium
is a minute more-or-less spherical
structure borne on a long stalk in the
shelter of each concave scale-like leaf on
the catkin-like male shoot.
ripe, the antheridia release the flagellated antherozoids, or male gametes, which swim across the
surface film of moisture, attracted to chemicals released by the egg cell (or possible associated
tissues). The perianth may assist this by holding water. In the ripe archegonium, the ventral canal
cell and neck canal cells break down into mucilage which takes in water and expands, opening the
flask-like archegonium ready for an antherozoid to swim down to the egg where fertilisation occurs.
The diploid zygote typically develops into the sporophyte.
grows out from the perianth as a stalk topped by spore capsule, which is usually darkly coloured
and glossy when mature and may be more-or-less spherical but is usually elongated. In some
forms, the young sporophyte begins its development completely enclosed in gametophyte tissue
which grows up from the stem to enclose the sporophyte, forming a marsupium. This protective
sheath of stem tissue may bear leaves or bracts and may replace a perianth in some forms which
lack this structure and may occur at the end of the shoot or hang down from the branch at right
angles and may be a sizeable structure.
Spore Dispersal
The wall of the spore capsule is two to several cell layers thick and the walls of the cells develop
bands of thickening. The spores are produced by meiosis and so are haploid. As the ripe
capsule dries out, these bands of thickenings, which are deposited in very specific patterns,
create stresses in the shrinking tissues which causes the capsule to rupture and to split into four
valves (above right). Bearing the capsule at the end of a stalk enables the spores to be released
above the boundary layer of still air, assisting dispersal. This is further assisted by the discharge
mechanism. In most leafy liverworts, the spores are discharged by a water rupture mechanism.
Interspersed with the spores are swollen, elongated cells called elaters. These clear, water-filled
elaters have a double spiral band of wall-thickenings on the inside of the cell wall. The elaters
may be free or anchored to the inside walls of the spore capsule (sporangium) in liverworts, but
are anchored at their base in this mechanism. As they dry out, the spiral thickenings try to
contract, like stretched springs eventually breaking the water column inside which cavitates as it
suddenly fills with pockets of gas and the elater instantaneously untwists, tearing itself free from
the sporangium wall, flying and rotating into the air, spinning off attached spores.
In the thalloid liverworts, Marchantia and Pellia (see: bryophytes introduction) each elater has a
single spiral thickening, which is much weaker and wriggle about as they slowly untwist upon
drying. This hygroscopic mechanism slowly fluffs up the spore mass which can then be more
easily dispersed by wind. In some leafy liverworts, such as Frullania, each elater spans the
sporangium and is firmly attached to the sporangium floor at one end and to the roof at the other.
As the four valves of the capsule bend back, as the capsule dries and opens, the elaters are
momentarily stretched and each has a single spiral of thickening and is essentially a stretched
spring in the wall of a water-filled tube. In less than a second the elaters are torn free at their
lower ends, which moves in an arc (as when a stretched spring is bent backwards and then
released from one end) flinging out the spores!
Spore production is prolific. An estimated 24 000 spores occur in each capsule of Lophocolea
cuspidata. Under suitable conditions a spore will germinate into a young gametophyte. The
pattern of sporeling development depends on species and, for example, the young
gametophyte may pass through either a filamentous stage, a disc-like stage or a ball-of-cells
stage. Eventually the three-sided apical cell is formed. This may generate at first a shoot with
juvenile leaves, which may be single cells or a row of cells, and then later shoots which appear
more adult-like, before the gametophyte finally puts out mature shoots.
Some leafy liverworts can also reproduce asexually by specialised means, apart from incidental
fragmentation or separation of parts due to death of stem segments, some have deciduous
propagules: branches, leaves or perianths (Gymnocolea inflata), which readily detach and
regenerate into new plants. In Frullania fragifolia, the large antical lobes of the bilobed leaves
detach, leaving only the smaller postical lobes. Some have more specialised asexual propagules
called gemmae, which are structures specialised only for reproduction, having no other specific
function in the adult plant. In Lophozia ventricosa, gemmae are produced at the tips of leaves,
which may bear clusters of gemmae along their margins, each gemma consists of one or two cells
with dense cytoplasm which detach easily.
Plagiochila (Featherwort) - an example of a leafy liverwort
Plagiochila is a beautiful leafy liverwort known as Featherwort. This specimen is from the British isles in a locale where two similar species occur: Plagiochila porelloides (Lesser Featherwort) and Plagiocila asplenioides (Greater Featherwort). These two species can be hard to distinguish. P. asplenioides is the larger of the two, forming shoots up to 12 cm long and 9 mm wide, with leaves up to 4.5 mm long. The smaller P. porellioides forms shoots up to 7 cm long, up to 6 mm wide and with leaves up to 3 mm long. Lesser Featherwort has frequent thread-like branches that bind adjacent shoots together to form a mat, whereas Greater Featherwort lacks threadlike branches.

A woodland population of Plagiochila asplenioides growing among mosses (in particular the large moss Plagiomnium undulatum) in winter. This site is also shared with Early purple Orchids (Orchis mascula), Bugle (Ajuga reptans) and Bluebells (Hyacinthoides non-scripta). which were just beginning to put up new leaves at this time.



This specimen was slightly over 7 cm in length (the photo above shows about 1 cm of shoot length) up to 9 mm wide and with leaves up to 4 mm or more in length and so is compatible with P. asplenioides. Furthermore, this specimen bore three branches that where similar to the main shoot. Plagiochila has greatly reduced amphigastria (underleaves), which are generally considered absent. this liverwort is succubous, meaning the back margin (nearest the apex in this case) lies beneath the front margin of the next leaf nearest the apex. The front margin folds back and extends down the stem, forming a line where it joins the stem (insertion line) that curves slightly and ends at about the midline of the stem.

As well as reproducing sexually, Plagiochila can propagate asexually by shedding leaves which then regenerate into new plants.

Above: bunches of rhizoids can be seen on the nodes of the underside of the stem. According to one source, the leafy portions of the stems of P. asplenioides lack rhizoids, but I have not yet verified this.

The leaves of both species usually have minute teeth along their margins,but these teeth are often too minute to see without a microscope, as in this specimen.

The perianth of Plagiochila, when it forms, consists of two modified leaf-like structures or valves and is laterally compressed.

Plagiochila porelloides is found on woodland banks, streamsides, and sheltered areas on upland slopes. P. asplenioides is the more common species in the south and east of Britain and is found in damp turf, woodlands, chalky slopes, fens, stream banks, hedgerows, and on rocks and rotting wood.
Watson, E.V. 1964. The Structure and life of Bryophytes. Hutchinson University Library.
http://rbg-web2.rbge.org.uk/bbs/Activities/liverworts/Diplophyllum%20albicans.pdf
http://rbg-web2.rbge.org.uk/bbs/Activities/liverworts/Frullania%20tamarisci.pdf
http://rbg-web2.rbge.org.uk/bbs/Activities/liverworts/Frullania%20fragilifolia.pdf
http://rbg-web2.rbge.org.uk/bbs/Activities/liverworts/Lophozia%20ventricosa.pdf
http://rbg-web2.rbge.org.uk/bbs/Activities/liverworts/Gymnocolea%20inflata.pdf
http://rbg-web2.rbge.org.uk/bbs/Activities/liverworts/Lophocolea%20bidentata.pdf
http://rbg-web2.rbge.org.uk/bbs/Activities/liverworts/Lophocolea%20heterophylla.pdf
Article created: 26 Nov 2016
Article updated: 21 Jan 2020