| |
|
|
|
|
|
| Bryophytes
- mosses |

Above:
epiphytes growing on the bark of an
oak tree; the grey-green encrusting epiphytes at the top of the
picture are lichens (a fungus host and an algal or cyanobacterial
endosymbiont) and lower down is a mat of epiphytic moss. The tip of
one of these mosses is shown magnified above. This moss is probably
a species of either Brachythecium or Isothecium, similar-looking mosses
that both grow as epiphytes. Characteristic of Brachythecium this moss has oval concave
leaves with distinctly pointed tips and small marginal teeth
(denticles) and midribs reaching to just above half-way along the
leaf.

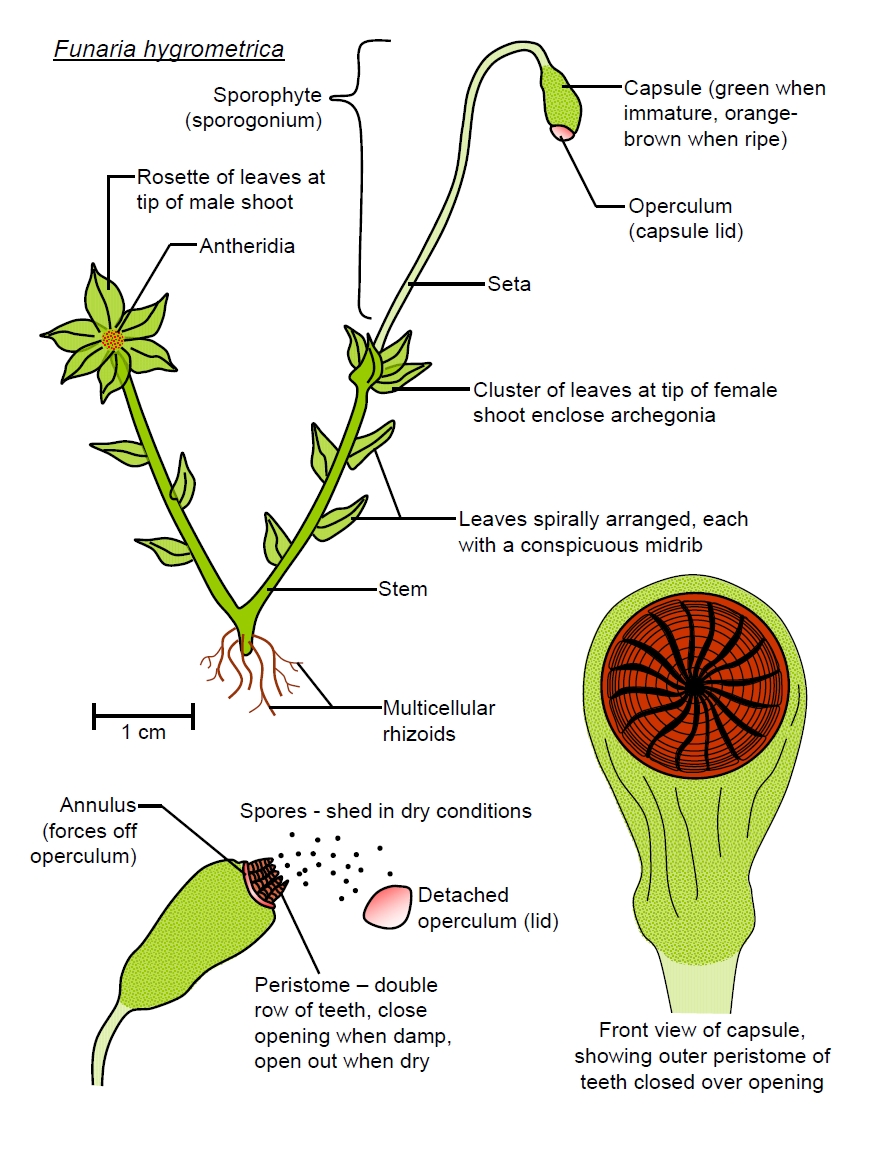
Funaria
hygrometrica is
a common moss found on bare soil in woodland, moorland and in
gardens and occurs frequently on burnt soil, such as at the sites of
recent bonfires.


Above: cross-section through a leaf of Funaria. Some mosses have more complicated midribs with various cells which transport materials, strengthen the midrib or have unknown functions. Mosses are bryophytes, along with liverworts and hornworts.

Above: Funaria hygrometrica growing on an old millstone.This moss usually colonises bare disturbed and nutrient-rich ground, such as old bonfire sites. The wavy seta is characteristic.
Bryophytes
- mosses and liverworts (including hornworts) are tiny plants
scarcely noticed by many, but closer inspection will reveal their
fantastic beauty and diversity of form. They are also informative
organisms in illustrating plant evolution and in illustrating how
physics can be beautifully exploited by some of the simpler
organisms. Liverworts, like Marchantia (see below), are small
plants with prostrate (lying flat against the substrate) leafy
thalli. (A thallus, plural thalli, is the name given to the body of
plants that lack definite roots, such as multicellular algae and
bryophytes). There are no roots, strictly speaking, but there are
rhizoids that serve for anchorage (but not water and mineral
absorption). The upper part of the thallus corresponds in some ways
to the green shoot-system of a tracheophyte (tracheophytes are
larger more complex plants, such as ferns, conifers and flowering
plants) in that the cells contain chloroplasts. In Marchantia these cells are arranged
in air chambers which contain branching columns of photosynthesising
cells which receive atmospheric carbon dioxide through stoma-like
pores (which are apparently incapable of closing like the stomata of
tracheophytes). A good place to find liverworts is clinging to the
soil of riverbanks.
Alternation
of Generations
The
leafy thallus of the bryophyte is haploid and produces gametes by
mitosis and is called the gametophyte. The spermatozoids are
biflagellate and swim through the surface film of moisture to reach
the eggs which are retained inside the thallus but open to the
outside via pores. The spermatozoids are attracted to chemicals
secreted by the egg and one can fertilise each egg. The result of
fertilisation is a diploid zygote which grows into a diploid plant
which produces spores by meiosis and is called the sporophyte. The
sporophyte remains attached to the parent gametophyte and is said to
be 'parasitic' on it since it absorbs much of its nutrition from the
gametophyte. However, it is not true parasitism since it is a mutual
symbiosis.
Water
Transport in Mosses
Mosses
differ from liverworts in having definite stems with (spiralling
arranged) leaves in addition to anchoring rhizoids.
Ectohydric
mosses
(a) rely mainly on water transport along the external surface of the
plant by capillarity. The spaces between the leaves and stem and
between papillae on the surface of leaves seem to be optimised to
act as capillary channels to draw water up and down the plant. The
rhizoids may form a dense felt-like tomentum and small leaf-like
structures called paraphyllia and both these structures can assist
capillarity. These mosses generally have very thin non-waxy cuticles
and are not waterproofed, allowing water to be absorbed along any
part of the body. these mosses dry out easily, but can tolerate
dryness, rehydrating when water returns and, contrary to popular
belief, they are not confined to damp habitats. Some mosses can
absorb fog and morning dew through their surfaces and so thrive on
arid mountains and in deserts. Most mosses are primarily ectohydric
and so have no specialised vascular tissue. Rather water can move
slowly from cell-to-cell across the cortex via plasmodesmata and
more readily via the apoplast, though internal movement in these
mosses is slow and external capillarity is rapid. Generally, the
upper tissues have the most rapid conduction and are supplied by
water first and some water may then move downwards inside the plant.
Ectohydric mosses will usually rehydrate within minutes when water
is added to dry plants.
Endohydric
mosses (b)
rely mostly on internal water transport and have vascular tissues.
They possess thin wax-like cuticles that confer some degree of water
repellency (though not to the extent seen in tracheophytes). They
have a primitive form of vascular tissue – the hydrome (consisting of hydroids and stereids) conducts water generally
down the stem and although non-lignified resembles xylem. The
hydroids are narrow elongated cells that join together to form
longitudinal tubes and are joined by slanted end-walls and resemble
tracheids. Like tracheids they lose their protoplasts when mature.
Hydroids also appear to contain lignin-like polyphenolic compounds
in their cell walls. However, hydroids differ from the tracheids and
vessel members of tracheophytes in that their end-walls are not
perforated by large pores to form pore/perforation plates (with few
exceptions), but instead contain many plasmodesmata; thus water
transport through these vessels will encounter more resistance and
be much slower than in tracheophyte xylem. The stereids are
cylindrical cells that connect together to form long thin fibers
with thick-walls and narrow lumens and probably have both a
mechanical supporting function and a water-conducting function.
(These cells are similar to the sclerenchyma cells of higher plants
but apparently do not contain lignin as sclerenchyma do). Thickened
cells in the cortex, especially the outer cortex also contribute to
support and may conduct water very slowly, probably mostly in a
radial direction, via the apoplast and through plasmodesmata
(symplast). The leptoids constitute the leptome
and resemble the phloem of tracheophytes and do indeed conduct
photoassimilates from sources to sinks. The leptome encloses the
hydrome. Like phloem tube elements these cells have degenerate
nuclei at maturity, but unlike phloem they lack highly porous sieve
plates, instead their end-walls contain numerous plasmodesmata, so
again conduction in the leptome is much slower than in phloem. True
leaf traces may be present – branches of vascular tissue that
connect the midribs of the leaves to the hydrome of the stem.
However, some species have pseudo leaf-traces – vascular strands
from the leaf midribs that end blindly in the cortex and do not
connect directly to the central strand of the stem.
Myxohydric
mosses
employ both the ectohydric and endohydric water-conducting pathways
in varying ratios. It can be argued that most, if not all, mosses
are really mixohhydric in that conduction of water occurs by both
external and internal routes to varying degrees, even when a central
strand is absent. Internal conduction may account for as little as
1% of water transport (ectohydric) to 70% or more (endohydric).
The diagrams below illustrate the structures of cross-sections
through the stems of an ectohydric moss (a) and an endohydric moss
(b).



Above and left: A transverse section through a moss identified as Polytrichum (an endohydric moss). The central strand with its hydroids is clearly visible. Typically in Polytrichum, however, there is a central strand of hydroids (hydrome) surrounded by a ring of leptoids (leptome). Some mosses have just the central strand of hydroids whilst others have only elongated parenchyma cells which transport food along the stem.

The
photomicrographs below show the structure of the moss pictured at
the top of this page.

The
tip of a single moss stem. These stems may be several cm long and
run mostly horizontally along the substratum (tree bark in this
instance) but the tip regions may stand upright.

The
leaves of this moss are sheets that are only one-cell thick. The
cells are elongate and packed with chloroplasts for photosynthesis.
The cells of moss leaves may be short and box-like, more-or-less
spherical or considerably elongated as in this moss. A useful
article on moss leaf variation is: https://www.disjunctnaturalists.com/articles1/mosses.htm.



Click photos to enlarge. Above and below: details of the marginal spines (denticles).

Moss
leaves may or may not have denticles, according to species. The
denticles may occur on the whole leaf margin or only towards the
apex. The denticles may be of this type, in which each
denticle is an extension of a peripheral cell or they may be part of
a distinct margin made up of specialised cells.

Below: close-up view of the midrib (commonly called a nerve by bryologists) which consists of cylinders of more elongated cells. Notice how the slanting end-walls of these cells are often tightly and closely pressed together - suggestive of water-conducting tissue. The midrib also provides the leaf with additional support.The leaves of some mosses lack a midrib, or it may only extend part way towards the leaf tip. The midrib may consist of a single strand of elongated cells or of multiple strands. It may be single or double.

Above: bundles of red-brown root-like rhizoids branch from the under-surface of the prostrate (horizontal or lying down) part of the stem. This moss is a pleurocarpous moss (a moss usually with prostrate habit / creeping stems that put out a number of upright setae bearing capsules, 'pleurocarpus' literally means 'lateral fruiting'). These anchor the moss plants in the tree bark, or in matter that has accumulated on the bark. They may have some role in water-transport by acting as wicks to draw water up along the outside of the rhizoids by capillarity, but they are probably not major organs of water absorption.
Many mosses are not pleurocarpos but acrocarpous: these are usually upright mosses (or prostrate turning upright at the ends) bearing a single terminal seta and capsule ('acrocarpous' literally means 'top fruiting').
Below: a section through the moss stem. The image is unfocused in places due to the thickness of the section which was simply cut with a scalpel and not sectioned on a microtome. More experienced bryologists have their own technique for cutting such sections with a razor blade. Nevertheless, the stem can be seen to comprise two regions - an outer cortex of smaller green thick-walled cells that give support to the stem (and probably carry out some photosynthesis) and an inner region of larger cells containing granules (starch grains and/or chloroplasts? This was a smallish stem, and the larger stems, though still tiny, can be surprisingly tough to cut.


It is hard to stay whether or not this moss has specialised water-conducting tissue; a longitudinal section would be needed to show this (there may be elongated parenchyma for food transport). Certainly when slicing the stems, the inner cells often frayed from inside their tough cortex, suggesting a filamentous nature. I have not tried to cut lengthwise with my scalpel, this would be tricky, but I might have a go. The greenish structures covering the surface of the stem are epiphytes - probably cyanobacteria, so epiphytes have epiphytes!
Left: a sector of cortex from the moss Polytrichum sp. showing the outer cortex of 4 or 5 cell layers of small and thick-walled cells which are clearly adapted to provide additional mechanical support. The middle cortex consists of larger cells, which like those of the outermost cortex, contain starch grains as a food reserve. The innermost cortex consists of large and angular parenchyma cells (assuming that these are not elongated cells in longitudinal section) with non-thickened walls. Each cell acts as a pressurised unit, inflating the stem when turgid and giving the plant mechanical support. Inside this is a central strand of conducting tissue (as illustrated above).

Below: a section through the cortex of Polytrichum sp. showing a leaf trace - a cluster of about 20 or so small cells which extends into a leaf higher up the axis, continuing along the midrib of the leaf. In gymnosperms and angiosperms such a trace would contain conducting tissue, such as xylem, in mosses, however, the leaf trace cells are less specialised and are hydroids in this case. In this cross-section there were 3 leaf traces in total. (Many bryophytes have three ranks of leaves).

Reproduction
in Bryophytes
Bryophytes
have extraordinary life-cycles in which the generations alternate
between tow genetically distinct forms - the haploid gametophyte,
which have already looked at and the diploid sporophyte. This is the
so-called alternation
of generations.
Haploid: Possessing only one set of chromosomes (like spermatozoa
and ova in humans).
Diploid: Possessing two sets of chromosomes - one maternal and one
paternal (like humans).
Curiously, the haploid form is the dominant growing stage and the
adult form in bryophytes. It produces gametes by mitosis (instead of
by meiosis as in diploid organisms) and so is the mature sexual
life-stage. The biflagellate
spermatozoids
(or antherozoids)are produced inside
containers called antheridia (singular antheridium),
from which they are released when ripe, and they use their two
flagella to swim toward the archegonium (plural archegonia), which
is typically a vase-like structure with an egg cell at the bottom of
it. The spermatozoids are attracted to the ova which releases
chemicals that the sperm sense. They will swim across in the film of
surface moisture covering clumps of bryophytes to reach her. One
spermatozoid only will fertilise an ovum, producing a diploid cell
or zygote. This cell then develops into the sporophyte whilst still
attached to the parent gametophyte. The sporophyte comprises an
elongated stalk or seta (plural setae) rooted in the gametophyte
tissue (at the base of what was the archegonium) by its 'foot'. The
free tip of the seta swells and develops into a spore-packed
capsule.
Spore
Dispersal Mechanisms
Bryophytes
have many ingenious mechanisms to assist in dispersal of their
spores.
Mosses. When mature, the spore
capsule dries and
typically a lid (operculum) detaches from the spore
capsule. Drying of the capsule is typically aided
by stomata-like pores which become uncovered when the calyptra is
shed and are generally located in the lower region of the capsul.
Once uncovered these stomata do not open or close but remain open
and air gradually replaces fluid in the intercellular spaces as the
capsule dries. The opening or mouth of and inner peristome when it
is double) of which there are often 16 teeth. These teeth have a
two-play structure and uneven thickenings in their cell walls, which
causes them to open or close with changes in humidity. In humid
conditions the teeth close, preventing spore discharge, whilst in
dry conditions the peristome opens, allowing spores to be discharged
and carried on the wind. (It is also possible that repeated
movements of the peristome can flick spores from the capsule). The
seta, typically one to a few cm in length, helps raise the capsule
above the boundary layer of stagnant air, to facilitate dispersal.
The seta may also twist rapidly one way and then the other as it
becomes moist or dries, which presumably aids dispersal by shaking
the spore capsule.
Liverworts. Liverwort spore capsules
lack peristomes. That of Pellia is spherical and when dry
splits into four valves which open to expose a mass of spores
intermingled with hair-like structures called elaters, some of which
are attached inside the capsule base and others are loose. The
elaters undergo writhing movements as they dry, dislodging the
spores for gradual dispersal.
Once the spore germinate they develop into new gametophytes.
Sporophyte
Spore Dispersal

Above: sporophyte of Mnium hornum (Swan's Neck Thyme Moss) emerging from an erect gametophyte, amid a patch of other gametophytes (also of Minum hornum). The sporophyte consists of a spore capsule atop a stalk or seta and the foot of the seta which draws nourishment from the leafy parental gametophyte to which it is attached. Take a closer look at Mnium hornum.
Below: a cross-section through the seta or stalk of a sporophyte. There are three distinct types of cells here - the small and very thick walled outer protective and supporting cells (orange) and the intermediate yellow cells (leptoids?) and the central strand of small cells (hydroids) which formed the conductive tissue which carried nutrients and water to the sporophyte tip as it was developing. Some mosses exceptionally have setae 5 to 10 cm in height.

Note the small thick-walled cells in the outer layers of the seta sectioned above. These are protective and supporting cells. Positioning tissue with the highest tensile strength towards the outside of the seta increases the stiffness of the seta, whereas placing these cells in the center would provide less support. The large parenchyma cells inside this tough outer sheath also have an important role in support when pressurized with water (sap in their vacuoles) contributing to stiffness by acting as a 'pressurized cellular solid' rather like the cells in polystyrene foam which are pressurized with air.

Above: Moss capsules which have shed their opercula caps removed to show the peristome teeth.

Above: a moss spore capsule with a beaked operculum. Note that the capsule is divisible into three general regions: the apex consists of the peristome (teeth) covered by the operculum (lid) which detaches when the capsule is ripe. The swelling at the base of the capsule, where the seta joins is the apophysis. The middle section contains the spore-forming mass surrounded by a jacket of photosynthetic tissue which contains air spaces supported by strands called trabeculae. (In some mosses the spore-forming mass extends into the seta so there is an apophysis as such). This type of beaked capsule is characteristic of the moss order Hypnales.

Above: the calyptra hood is still in place on this Kindbergia capsule, covering the operculum lid.

Above:capsules in various stages of development. This moss was found growing at the base of a black poplar tree, and other specimens around the base of an oak tree.



The 'perfect' peristome consists of 16 outer teeth (the exostome) as in this case which alternate with 16 inner teeth (the endostome) with a group of 3 or 4 hairlike cilia in between each pair of adjacent inner teeth. In many mosses, however, one or more of these components is reduced or absent and some have no peristome at all.

The peristome is hygroscopic and opens in dry conditions to allow spore dispersal. The outer teeth are extremely sensitive to humidity and can sometimes be seen to rapidly pulse in and out, in synchrony, under the microscope.

Although the slightest disturbance of a moss capsule, causing the seta to vibrate, will easily disperse the spores when the peristome is open, the rhythmic opening and closing of the peristome with changes in humidity, also flicks spores out from the capsule. This latter form of dispersal may be particularly important because mosses are small and so barely break through the boundary layer of still air to reach the turbulent air layer where spores can be readily carried away by the wind. In the picture above, a spore can be seen attached to the peristome and may be easily flicked away as teh peristome opens.

Above and below: spores from the Brachythecium-type moss.

This epiphyte had leaves without a prominent midrib but with more elongated cells forming a weak nerve which extended only about half-way towards the tip. The leaf tips were drawn out into quite long points made up of leaf cells:



The leaves of this moss lack midribs and are extremely tapered and pointed. The margins are smooth (or sometimes with very tiny denticle-like projections more obvious in some specimens) and they frequently had some brown-red cells perhaps containing anthocyanin. These leaf features are typical of Brachythecium and some related genera.

The
sporophyte derives some of its nutrition from the parent
gametophyte, via its foot and is said to be 'parasitic' on the
gametophyte. However, the sporophyte is capable of photosynthesis
when young and green, and so produces some of its own food. (Also,
this isn't true parasitism, since the gametophyte is really
investing in its offspring). There is a small space between the edge
of the foot and the adjacent cells of the gametophyte and the
gametophyte cells are thought to actively pump nutrients into this
space and may be modified into transfer
cells
(that is they have wall invaginations to increase the surface area
of their ell-surface membrane to accommodate more pumps). The foot
absorbs these nutrients (and may also have transfer cells to pump
the nutrients in) and these are then conducted along the seta (which
has leptoid and hydroids in at least some cases) to the developing
cells. The seta and then later the spore capsule typically turn from
green to red-brown in mosses as the cells die and dehydrate during
spore release. The tip of the developing sporophyte is typically
covered in a sheath or calyptra, which is the remains of the neck
and venter of the flask-shaped archegonium in which the zygote
germinates.



Above: the tips of developing sporophytes, with the calyptra removed. These sporophyte tips were originally covered in the calyptra sheath, as shown below left, and detaching (below right).
Asexual reproduction. Both mosses and
liverworts have a variety of means of reproducing asexually. Mosses
may regenerate from broken or fragmented parts, including single
leaves. specialised structures, called gemmae (singular gemma) are
multicellular balls, ellipsoids or discs which are easily detached
and which produce new gametophyte plants on germination. Gemmules
may be born on any part of the plant. In some liverworts they are
borne in pretty cup-like structures.
In
the moss, whose sporophyte is shown in the photographs above, the
adxaial (upper) surfaces of many of the leaves carried spherical
balls of 2 or 4 cells each, with up to 3 found on any single
leaf. These are unlikely to be gemmae, however, as they lack
the degree of organisation typical of moss gemmae and are enclosed
in a thick slime capsule. This is probably a trebouxioid alga
growing epiphytically upon the moss or on the substrate shared with
the moss. These balls of cells are shown below. Some cells within
these balls, which were quite frequent on this moss, are evidently
dividing within their common slime envelope.



Below: cells in the leaf of an unidentified species of moss, with large chloroplasts. The apoplast system (cell walls and the middle lamella which glues neighbouring cells together) provides both support and acts as a conduit for water movement through the leaf. Right, elongated cells of the leaf midrib provide mechanical support and transport photoassimilates from cell to cell, via plasmodesmata, in the symplast transport pathway.


The sporophytes below suggest that this moss is Brachythecium
rutabulum (the Rough-Stalked Feather-moss) with tiny
protuberances on the seta giving it a rough appearance.

Below: a type of moss with specialised borders to its leaf margins made up of specialized cells. This type of border occurs in a number of mosses, including the common Mnium hornum, the Swan's-neck Thyme-moss.

Below: The leaves of some mosses bear rows of cells called lamellae, either on the whole leaf surface, as here, or on the midrib/nerve. This leaf, seen in section, belongs to the moss Polytrichum.


Above and below: one of the anatomically most sophisticated moss leaves is that of Polytrichum. The photosynthetic cells are arranged in vertical longitudinal sheets, each one cell wide (seen in transverse or cross-section in these diagrams) on the upper surface of each leaf. Most of the chloroplasts occur in these cells. The outermost end or terminal cell of each lamella has a characteristic notch. These notches are thought to form capillary channels for the conductance of an external water film, whilst the spaces between the lamellae presumably remain dry for gas exchange during photosynthesis. Internally, the supporting rib or costa of each leaf has a high degree of differentiation into different cell and tissue types: thick-walled stereids provide mechanical support in mature leaves and can also transport water through the apoplast (their cell walls which are porous and act like a sponge drawing in water by capillary action). There is an internal layer of large conducting cell, some authors label these as hydroids, but I have recently (25/2/2025) referred to Charles Hébant's paper on the related moss Dawsonia in which these cells are classed as food-conducting parenchyma cells (essentially leptoids) that transport photoassimilate (organic products made in the leaf by photosynthesis). Thus, I have updated the labels to agree with Hébant (1976), though there can be variations in different moss species, hydroids are cells that lack protoplasts, however, this is not easy to tell in this section of fixed material. A thick-walled layer of epidermal cells covers the external surface of the back of the leaf.
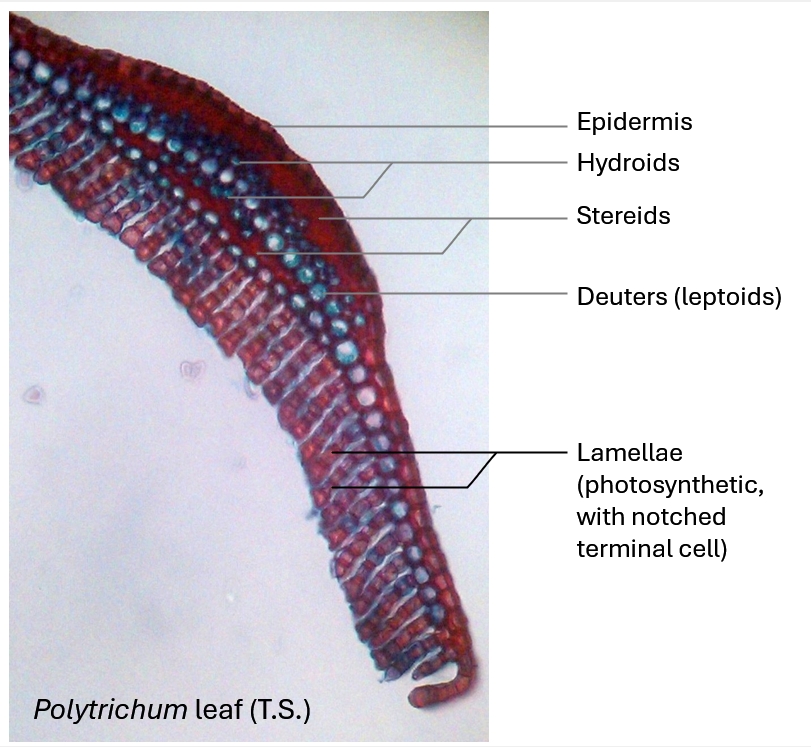
The large cells are probably deuters and the smaller cells hydroids.
More Bryophyte Pages:


Article updated:
29 Oct 2016
10 Nov 2016
27 Nov 2016
18 Feb 2017
12 Aug 2018
19 Dec 2019
24 Dec 2019
27 Feb 2022
08 Feb 2025
15 Feb 2025
25 Feb 2025