








Millipedes belong to a group of arthropods (invertebrates with jointed limbs) called the myriapods ('many
legs') which includes four extant divisions: the millipedes (diplopods), the centipedes (chilopods), the
symphylans and the pauropods. Like all artghropods, they are encased in an exoskeleton of cuticle.
Millipedes are the largest land-dwelling arthropods, the African giant millipedes (chongololo) may reach
32 to 38.5 cm in length and 6.7 cm in circumference (for Archispirostreptus gigas). Some prehistoric
millipedes, however, reached much larger proportions: Arthopleura was up to 2.3 m long, with a broad
and flattened body. Millipedes were amongst the first animals to colonise land in the Silurian, some 400
million years ago. Millipedes may live up to 10 years. Millipedes vary enormously in form and colour and
there are about 8000 recognised species. Some millipedes are bioluminescent, possessing a light
emitting protein beneath their cuticles, which generates a greenish or blueish light.
Millipede Bauplan
The first body segment behind the head is called the collum and this collar-like neck bears no legs.
Behind the collum are several haplosegments (often 3, i.e. segments 2, 3 and 4, as in this model) which
bear a single pair of legs and often a single pair of spiracles. A 'haplosegment' is a normal single
metameric segment, that is it contains a single set of some of the organ systems within it, such as a single
pair of respiratory spiracles and a single pair of lateral nerves emanating from the double ventral
nerve cord. This is distinguished from the majority of body segments in millipedes, which are double
segments or diplosegments. Each diplosegment may appear externally as a single segment but is
actually the fusion of two body segments. Therefore, diplosegments have two copies of some of the
organ systems: two pairs of spiracles and two pairs of cordal (segmental) nerves and two pairs of legs.
The legs of the haplosegments, especially the first haplosegment, are often reduced in size.
The third segment is also called the genital segment or gonosegment, since in males it contains either
one pair (sometimes two) of penes on or near the basal segments (coxae, singular = coxa) of the legs or
a single penis on the undersurface in a groove between the bases of the legs. In females the
gonosegment has a pair of lateral slits, each of which opens into a pouch-like atrium, called a vulva,
which can be protracted to receive sperm from the male during copulation, or retracted and covered by a
sclerotised (hardened) hood. Received sperm are passed to a seminal receptacle for storage: the
female will fertilise the eggs as they are laid later.
The penis or penes do not function as intromittent organs, however. Instead the seventh segment of
males has one or both pairs of legs modified (or one pair plus one pair on the 8th segment) into
gonopods. In some forms, the first pair of gonopods serve as protective shields for the second pair
which are the intromittent organs. Our model has one pair of gonopods on the seventh segment. During
mating, the penis or penes pass sperm to the gonpodium (the male folds the front of his body over to
bring the penis in contact with a gonopod) which have a series of canals and reservoirs to hold the
sperm. The gonopods will then inject the sperm into the protracted vulvae of the female. The gonopods
have complex and species-specific morphology - this ensures that only gonopods from a male of the
same species will fit inside the female. For this reason, gonopod morphology (and likewise for the
intromittent organs of insects) are important in the study of millipede evolution.
The pair of testes are tubular organs, connected together by transverse connecting tubules, and extend
backwards from the second segment (to the 40th in millipedes with that many segments) and is situated
In females, one pair of fused elongated and tubular ovaries is situated between the (mid)gut and ventral
nerve cord in a similar position to the testes in the male.
Each spiracle(or stigma, plural = stigmata) is located ventrally, near to and often just in front of each leg
base and opens into a tracheal pouch or vestibule from which emanate a number of tracheal tubes,
carrying air, and equipped with spiral thickenings of cuticle to prevent their collapse. Some of the
tracheae are bundled into two longitudinal ventrolateral (towards the sides and underside); other bundles
connect opposite pairs of pouches together and ventral transverse bundles connect the two longitudinal
trunks together. As in insects, these ultimately end in branches of fine tracheoles, which supply individual
cells and groups of cells with oxygen and remove carbon dioxide. As well as relying on diffusion of air
within the tracheoles, for gas exchange, movements of the body and legs possibly assist air flow as they
do in insects.
legs') which includes four extant divisions: the millipedes (diplopods), the centipedes (chilopods), the
symphylans and the pauropods. Like all artghropods, they are encased in an exoskeleton of cuticle.
Millipedes are the largest land-dwelling arthropods, the African giant millipedes (chongololo) may reach
32 to 38.5 cm in length and 6.7 cm in circumference (for Archispirostreptus gigas). Some prehistoric
millipedes, however, reached much larger proportions: Arthopleura was up to 2.3 m long, with a broad
and flattened body. Millipedes were amongst the first animals to colonise land in the Silurian, some 400
million years ago. Millipedes may live up to 10 years. Millipedes vary enormously in form and colour and
there are about 8000 recognised species. Some millipedes are bioluminescent, possessing a light
emitting protein beneath their cuticles, which generates a greenish or blueish light.
Millipede Bauplan
The first body segment behind the head is called the collum and this collar-like neck bears no legs.
Behind the collum are several haplosegments (often 3, i.e. segments 2, 3 and 4, as in this model) which
bear a single pair of legs and often a single pair of spiracles. A 'haplosegment' is a normal single
metameric segment, that is it contains a single set of some of the organ systems within it, such as a single
pair of respiratory spiracles and a single pair of lateral nerves emanating from the double ventral
nerve cord. This is distinguished from the majority of body segments in millipedes, which are double
segments or diplosegments. Each diplosegment may appear externally as a single segment but is
actually the fusion of two body segments. Therefore, diplosegments have two copies of some of the
organ systems: two pairs of spiracles and two pairs of cordal (segmental) nerves and two pairs of legs.
The legs of the haplosegments, especially the first haplosegment, are often reduced in size.
The third segment is also called the genital segment or gonosegment, since in males it contains either
one pair (sometimes two) of penes on or near the basal segments (coxae, singular = coxa) of the legs or
a single penis on the undersurface in a groove between the bases of the legs. In females the
gonosegment has a pair of lateral slits, each of which opens into a pouch-like atrium, called a vulva,
which can be protracted to receive sperm from the male during copulation, or retracted and covered by a
sclerotised (hardened) hood. Received sperm are passed to a seminal receptacle for storage: the
female will fertilise the eggs as they are laid later.
The penis or penes do not function as intromittent organs, however. Instead the seventh segment of
males has one or both pairs of legs modified (or one pair plus one pair on the 8th segment) into
gonopods. In some forms, the first pair of gonopods serve as protective shields for the second pair
which are the intromittent organs. Our model has one pair of gonopods on the seventh segment. During
mating, the penis or penes pass sperm to the gonpodium (the male folds the front of his body over to
bring the penis in contact with a gonopod) which have a series of canals and reservoirs to hold the
sperm. The gonopods will then inject the sperm into the protracted vulvae of the female. The gonopods
have complex and species-specific morphology - this ensures that only gonopods from a male of the
same species will fit inside the female. For this reason, gonopod morphology (and likewise for the
intromittent organs of insects) are important in the study of millipede evolution.
The pair of testes are tubular organs, connected together by transverse connecting tubules, and extend
backwards from the second segment (to the 40th in millipedes with that many segments) and is situated
In females, one pair of fused elongated and tubular ovaries is situated between the (mid)gut and ventral
nerve cord in a similar position to the testes in the male.
Each spiracle(or stigma, plural = stigmata) is located ventrally, near to and often just in front of each leg
base and opens into a tracheal pouch or vestibule from which emanate a number of tracheal tubes,
carrying air, and equipped with spiral thickenings of cuticle to prevent their collapse. Some of the
tracheae are bundled into two longitudinal ventrolateral (towards the sides and underside); other bundles
connect opposite pairs of pouches together and ventral transverse bundles connect the two longitudinal
trunks together. As in insects, these ultimately end in branches of fine tracheoles, which supply individual
cells and groups of cells with oxygen and remove carbon dioxide. As well as relying on diffusion of air
within the tracheoles, for gas exchange, movements of the body and legs possibly assist air flow as they
do in insects.
| | | | | | | | |
| Millipedes (Diplopoda) |
Above: legs removed from one side to show spiracles. Unlike those of many insects, the
spiracles of millipedes can not be opened and closed, but instead are permanently open.
Millipedes have an open circulatory system with haemolymph bathing tissues directly. The
haemolymph contains the respiratory pigment, haemocyanin (Hc) in some species. This blue
pigment contains copper rather than iron which gives haemoglobin (Hb) its red colour in the
blood of vertebrates. Whereas mammalian Hb molecules are proteins composed of four
polypeptide subunits, millipede Hc molecules have 36 subunits, arranged in 6 groups of six.
The haemolymph is circulated, as in insects, by a tubular dorsal heart (dorsal vessel) which
collects haemolymph via two pairs of pores, or ostia, in each diplosegment (as opposed to one
pair per segment in insects) and peristaltic waves of muscular contraction pump the
haemolymph to the anterior end, where the dorsal vessel is drawn out as a cephalic artery or
into a pair of short dorsal aorta (lacking ostia) which send out arteries to supply the head. The
posterior end of the dorsal vessel ends blindly. Some blood may flow back along a median
ventral blood vessel which may send out other branches, eventually blood leaves the open
ends of the arteries and enters the haemocoel to become haemolymph which bathes the
organs directly.
The gut, as in insects, is divided into foregut, midgut and hindgut. The hindgut opens as the
anus, which is situated ventrally on the end-most segment or telson. The telson (or the
segment immediately before it, it's hard to tell) is legless and consists of a perianal ring of
cuticle, a dorsal cuticular extension of which, called the epiproct forms a roof over the anus,
whilst a ventral plate or hypoproct encloses it from beneath and two side plates or paraprocts
partially enclose it from the sides. These plates are visible in our model.
One or two pairs of excretory organs, Malpighian tubules, emerge from the midgut / hindgut
boundary and loop back and forth a couple of times, extending anteriorly to the oesophagus
(part of the foregut) and posteriorly to the rectum. Their ends are blind and these tubules are
bathed in haemolymph from which they extract nitrogenous wastes, mostly ammonia, but some
uric acid as well. This waste is then passed to the gut to be voided with the faeces. Uric acid is
also deposited in the cuticle and shed with each moult. Tracheoles cover the outside of each
Malpighian tubule, supplying this energy-consuming organ with oxygen.
spiracles of millipedes can not be opened and closed, but instead are permanently open.
Millipedes have an open circulatory system with haemolymph bathing tissues directly. The
haemolymph contains the respiratory pigment, haemocyanin (Hc) in some species. This blue
pigment contains copper rather than iron which gives haemoglobin (Hb) its red colour in the
blood of vertebrates. Whereas mammalian Hb molecules are proteins composed of four
polypeptide subunits, millipede Hc molecules have 36 subunits, arranged in 6 groups of six.
The haemolymph is circulated, as in insects, by a tubular dorsal heart (dorsal vessel) which
collects haemolymph via two pairs of pores, or ostia, in each diplosegment (as opposed to one
pair per segment in insects) and peristaltic waves of muscular contraction pump the
haemolymph to the anterior end, where the dorsal vessel is drawn out as a cephalic artery or
into a pair of short dorsal aorta (lacking ostia) which send out arteries to supply the head. The
posterior end of the dorsal vessel ends blindly. Some blood may flow back along a median
ventral blood vessel which may send out other branches, eventually blood leaves the open
ends of the arteries and enters the haemocoel to become haemolymph which bathes the
organs directly.
The gut, as in insects, is divided into foregut, midgut and hindgut. The hindgut opens as the
anus, which is situated ventrally on the end-most segment or telson. The telson (or the
segment immediately before it, it's hard to tell) is legless and consists of a perianal ring of
cuticle, a dorsal cuticular extension of which, called the epiproct forms a roof over the anus,
whilst a ventral plate or hypoproct encloses it from beneath and two side plates or paraprocts
partially enclose it from the sides. These plates are visible in our model.
One or two pairs of excretory organs, Malpighian tubules, emerge from the midgut / hindgut
boundary and loop back and forth a couple of times, extending anteriorly to the oesophagus
(part of the foregut) and posteriorly to the rectum. Their ends are blind and these tubules are
bathed in haemolymph from which they extract nitrogenous wastes, mostly ammonia, but some
uric acid as well. This waste is then passed to the gut to be voided with the faeces. Uric acid is
also deposited in the cuticle and shed with each moult. Tracheoles cover the outside of each
Malpighian tubule, supplying this energy-consuming organ with oxygen.
Each segment is encased in an exoskeleton of cuticle reinforced with calcium carbonate. The
back (dorsum) is protected by a thick dorsal plate or tergite, the sides by the pleura and on
the underside the sternal plates support the legs. On or near to the bases of the tergites
some species have openings to repugnatorial glands, which may be lacking on the first few
segments. When threatened, millipedes with repugnatorial glands may squirt fluid up to 30 cm
from these openings. This liquid is an irritant containing different chemicals in different
taxonomic groups, including: benzoquinones, alkaloids, phenols, hydrogen cyanide,
nitroalkanes, cyanogenic compounds and terpenoids. This liquid can cause scarring to the
eyes and inner lining of the mouth and so one should be careful when handling millipedes.
Some millipedes can also protect themselves by coiling up or rolling into a ball (pill millipedes)
to protect their more vulnerable parts.
back (dorsum) is protected by a thick dorsal plate or tergite, the sides by the pleura and on
the underside the sternal plates support the legs. On or near to the bases of the tergites
some species have openings to repugnatorial glands, which may be lacking on the first few
segments. When threatened, millipedes with repugnatorial glands may squirt fluid up to 30 cm
from these openings. This liquid is an irritant containing different chemicals in different
taxonomic groups, including: benzoquinones, alkaloids, phenols, hydrogen cyanide,
nitroalkanes, cyanogenic compounds and terpenoids. This liquid can cause scarring to the
eyes and inner lining of the mouth and so one should be careful when handling millipedes.
Some millipedes can also protect themselves by coiling up or rolling into a ball (pill millipedes)
to protect their more vulnerable parts.
Above: openings (ozopores) of the repugnatorial glands.
Above: the head of our millipede model, viewed face-on. The eyes are fields of 2 to 80 simple
eyes or ocelli (singular = ocellus) which primarily function in detecting the direction of a light
source and probably movement of shadows but are thought not to do much in the way of image
formation. Some millipedes, for example those living in caves (troglomorphic forms) are blind. A
pair of antennae emerge from the head. Each antenna is made up of 8 antennomeres
('segments'): the basal most scape, then the pedicel and then 6 flagellomeres (F1 most basally
to F6 at the apex) making up the flagellum. Most of the antennomeres contain touch-sensitive
hairs (trichoid and chaetiform sensilla. several different types of tiny cone-like hairs, or
basiconic sensilla, most of which are probably olfactory receptors (as in insects) occur, often
grouped into strategically positioned fields, on F3, F4 and F5. The very short terminal F6
antennomere typically bears four large cones, arranged in a square, each with a pore at its
apex, which are probably gustatory (detecting chemicals on contact) and the antennae
constantly probe the surface as the animal moves. F6 is so short that the antenna appears to
be comprised of only 7 segments except when seen up close.
Positioned near the base of each antenna, somewhere between each antenna and eye, is
located an organ of Tomosvary, so there is one pair of these organs in total. These are pores
opening into pits, lined by cuticle, which contain sensory nerve endings embedded in the cuticle
covering the floor of the pit. Other myriapods have these organs, though their precise form is
very variable. For example, in the symphylan Scutigerella immaculata, the sensory cells put out
a reticulum of anastomosing cilia (Tomosvary, cited in: Bulletin of the British Myriapod and
isopod group, Volume 19 2003). The exact function of these organs is not known, however,
some have the appearance of mechanoreceptors (perhaps proprioceptors measuring
deformation of the cuticle) whilst others could be olfactory receptors or thermohygroreceptors.
One study on centipedes (Yamana et al., 1986: J. exp. Biol. 126, 297-314) found that the
organs were highly responsive to variations in carbon dioxide concentration and possibly pH
(whether this is physiological or not is uncertain). Others have suggested that these organs are
organs of equilibrium - sensing changes in direction of the millipede head.
The mouthparts consist of the upper lip or labrum, with a lower lip formed by fusion of the pair
of primary maxillae (no secondary maxillae occur in millipedes) to form the gnathochilarium
which is also equipped with basiconic sensilla (presumably chemoreceptive). This structure
bears 4 small sensory palps on its end. The sides of the mouth are covered by a pair of
mandibles, consisting of a basal segment (cardo) hinged with a distal segment (stipes) Most
millipedes feed on rotting vegetation, though some will graze seedlings, and others ingest soil
like earthworms. Some have piercing mouthparts to feed on plant juices and some are
omnivorous or carnivorous, preying upon centipedes, earthworms, insects, etc. The long
midgut secretes a peritrophic membrane which surrounds and packages the faeces (as in
insects).
The millipede brain
The millipede brain is morphologically similar to the insect brain consisting of a
supraoesophageal ganglion (situated above the oseophagus of the foregut), connected to the
sub-oesophageal ganglion (situated below the oesophagus) by a pair of circumoesophageal
commissures. This gives rise to the elongated ventral nerve cord which gives of segmental
nerves (two pairs per diplosegment). Like the insect brain, the brains of some millipedes can be
quite sophisticated, as evidences by their reproductive behaviour which is well studied in
juliform millipedes. Mating usually starts when the male communicates intent by antennal
tapping or head drumming or stridulating. The male usually climbs on to the back of the female,
clinging by means of special leg pads. Pheromones may also be involved. The male twists and
stretches his body until his gonopods, charged with sperm, lie opposite the vulvae of the
female, whilst holding the female with his legs. Each gonopod has a crescent-shaped tip,
composed of the longer telopodite and shorter endopodite. Sperm from the tip of the telopodite
is passed into the vulva, where it is stored by the female to fertilise the eggs as they are laid.
Between 10 and 300 eggs are laid in clusters in soil or humus. Narceus regurgitates material
which it moulds into a cup with its head and anterior legs. A single egg is laid into this cup,
which is then closed, sealed and polished. The finished capsule is then deposited in humus or
a safe crevice and will be eaten by the hatchling. Glomerus (the European pill millipede) forms
a similar capsule from excrement. As well as providing protection for the developing egg and
food for the hatchling, these capsules possibly transmit microbial gut symbionts to the offspring.
Some forms build a nest from excrement, consisting of a thin-walled and domed chamber with a
chimney. Eggs are deposited inside via the chimney, which is then sealed and covered with
grass and detritus to conceal it from predators. Either the female or male, in some species, may
lie coiled about the nest for several weeks, protecting the eggs. Some build nests in soil,
excavating chambers whose inner walls are reinforced with excrement, yet others enclose their
eggs in silk cocoons.
The hatchling is a legless pupoid which moults into a juvenile with 3 pairs of legs (like an insect)
but more segments and legs are added at each molt. This phenomenon of a juvenile lacking
the full complement of adult segments, with more (1-4) being added at each molt, is called
anamorphosis. (Insect juveniles have the full adult complement of segments, a developmental
strategy called epimorphosis). Some forms molt within the safety of molting chambers, similar to
egg nests. Some tropical forms will survive the dry season encased in such a chamber.
Millipedes usually eat their molted exoskeleton, perhaps recycling the calcium with it.
Parthenogenesis may occur in certain millipedes in which males are rare.
Locomotion
Millipedes are so-called because they have many legs, though not quite a thousand as the
name suggests, though one species of threadlike vermiform (wormlike) millipede has 750 legs.
Each leg moves backwards, whilst in contact with the substrate, during the power stroke and
then lifts up and moves forwards to reposition itself during the recovery stroke. As the power
stroke takes longer to complete more legs are on the ground at any one time, generating more
thrust. This is important since most millipedes plough through leaf litter and soil, searching for
food, which requires strength rather than speed. The bull-dozer like head helps them push their
way through and the calcium in their exoskeletons makes them stiff and rigid in these forms.
Short legs, often held vertically downwards, facilitate these ploughing movements. Shorter limbs
are slower but stronger, all other things being equal, but carnivorous millipedes and centipedes
have longer legs (very long in some centipedes) designed more for speed rather than strength.
In centipedes, serpentine undulations of the body may also occur in centipedes, but these are
considered disadvantageous as they reduce speed and some centipedes are designed to
reduced this tendency, e.g. with interlocking tergites. The legs operate in waves of activity as
can be clearly seen in his animation (external link) of a bioluminescent millipede walking:
National Geographic: glowing millipede
Note also the palpitating motion of the antennae and that the wavelength of leg movement is
about half the body length in this millipede.
eyes or ocelli (singular = ocellus) which primarily function in detecting the direction of a light
source and probably movement of shadows but are thought not to do much in the way of image
formation. Some millipedes, for example those living in caves (troglomorphic forms) are blind. A
pair of antennae emerge from the head. Each antenna is made up of 8 antennomeres
('segments'): the basal most scape, then the pedicel and then 6 flagellomeres (F1 most basally
to F6 at the apex) making up the flagellum. Most of the antennomeres contain touch-sensitive
hairs (trichoid and chaetiform sensilla. several different types of tiny cone-like hairs, or
basiconic sensilla, most of which are probably olfactory receptors (as in insects) occur, often
grouped into strategically positioned fields, on F3, F4 and F5. The very short terminal F6
antennomere typically bears four large cones, arranged in a square, each with a pore at its
apex, which are probably gustatory (detecting chemicals on contact) and the antennae
constantly probe the surface as the animal moves. F6 is so short that the antenna appears to
be comprised of only 7 segments except when seen up close.
Positioned near the base of each antenna, somewhere between each antenna and eye, is
located an organ of Tomosvary, so there is one pair of these organs in total. These are pores
opening into pits, lined by cuticle, which contain sensory nerve endings embedded in the cuticle
covering the floor of the pit. Other myriapods have these organs, though their precise form is
very variable. For example, in the symphylan Scutigerella immaculata, the sensory cells put out
a reticulum of anastomosing cilia (Tomosvary, cited in: Bulletin of the British Myriapod and
isopod group, Volume 19 2003). The exact function of these organs is not known, however,
some have the appearance of mechanoreceptors (perhaps proprioceptors measuring
deformation of the cuticle) whilst others could be olfactory receptors or thermohygroreceptors.
One study on centipedes (Yamana et al., 1986: J. exp. Biol. 126, 297-314) found that the
organs were highly responsive to variations in carbon dioxide concentration and possibly pH
(whether this is physiological or not is uncertain). Others have suggested that these organs are
organs of equilibrium - sensing changes in direction of the millipede head.
The mouthparts consist of the upper lip or labrum, with a lower lip formed by fusion of the pair
of primary maxillae (no secondary maxillae occur in millipedes) to form the gnathochilarium
which is also equipped with basiconic sensilla (presumably chemoreceptive). This structure
bears 4 small sensory palps on its end. The sides of the mouth are covered by a pair of
mandibles, consisting of a basal segment (cardo) hinged with a distal segment (stipes) Most
millipedes feed on rotting vegetation, though some will graze seedlings, and others ingest soil
like earthworms. Some have piercing mouthparts to feed on plant juices and some are
omnivorous or carnivorous, preying upon centipedes, earthworms, insects, etc. The long
midgut secretes a peritrophic membrane which surrounds and packages the faeces (as in
insects).
The millipede brain
The millipede brain is morphologically similar to the insect brain consisting of a
supraoesophageal ganglion (situated above the oseophagus of the foregut), connected to the
sub-oesophageal ganglion (situated below the oesophagus) by a pair of circumoesophageal
commissures. This gives rise to the elongated ventral nerve cord which gives of segmental
nerves (two pairs per diplosegment). Like the insect brain, the brains of some millipedes can be
quite sophisticated, as evidences by their reproductive behaviour which is well studied in
juliform millipedes. Mating usually starts when the male communicates intent by antennal
tapping or head drumming or stridulating. The male usually climbs on to the back of the female,
clinging by means of special leg pads. Pheromones may also be involved. The male twists and
stretches his body until his gonopods, charged with sperm, lie opposite the vulvae of the
female, whilst holding the female with his legs. Each gonopod has a crescent-shaped tip,
composed of the longer telopodite and shorter endopodite. Sperm from the tip of the telopodite
is passed into the vulva, where it is stored by the female to fertilise the eggs as they are laid.
Between 10 and 300 eggs are laid in clusters in soil or humus. Narceus regurgitates material
which it moulds into a cup with its head and anterior legs. A single egg is laid into this cup,
which is then closed, sealed and polished. The finished capsule is then deposited in humus or
a safe crevice and will be eaten by the hatchling. Glomerus (the European pill millipede) forms
a similar capsule from excrement. As well as providing protection for the developing egg and
food for the hatchling, these capsules possibly transmit microbial gut symbionts to the offspring.
Some forms build a nest from excrement, consisting of a thin-walled and domed chamber with a
chimney. Eggs are deposited inside via the chimney, which is then sealed and covered with
grass and detritus to conceal it from predators. Either the female or male, in some species, may
lie coiled about the nest for several weeks, protecting the eggs. Some build nests in soil,
excavating chambers whose inner walls are reinforced with excrement, yet others enclose their
eggs in silk cocoons.
The hatchling is a legless pupoid which moults into a juvenile with 3 pairs of legs (like an insect)
but more segments and legs are added at each molt. This phenomenon of a juvenile lacking
the full complement of adult segments, with more (1-4) being added at each molt, is called
anamorphosis. (Insect juveniles have the full adult complement of segments, a developmental
strategy called epimorphosis). Some forms molt within the safety of molting chambers, similar to
egg nests. Some tropical forms will survive the dry season encased in such a chamber.
Millipedes usually eat their molted exoskeleton, perhaps recycling the calcium with it.
Parthenogenesis may occur in certain millipedes in which males are rare.
Locomotion
Millipedes are so-called because they have many legs, though not quite a thousand as the
name suggests, though one species of threadlike vermiform (wormlike) millipede has 750 legs.
Each leg moves backwards, whilst in contact with the substrate, during the power stroke and
then lifts up and moves forwards to reposition itself during the recovery stroke. As the power
stroke takes longer to complete more legs are on the ground at any one time, generating more
thrust. This is important since most millipedes plough through leaf litter and soil, searching for
food, which requires strength rather than speed. The bull-dozer like head helps them push their
way through and the calcium in their exoskeletons makes them stiff and rigid in these forms.
Short legs, often held vertically downwards, facilitate these ploughing movements. Shorter limbs
are slower but stronger, all other things being equal, but carnivorous millipedes and centipedes
have longer legs (very long in some centipedes) designed more for speed rather than strength.
In centipedes, serpentine undulations of the body may also occur in centipedes, but these are
considered disadvantageous as they reduce speed and some centipedes are designed to
reduced this tendency, e.g. with interlocking tergites. The legs operate in waves of activity as
can be clearly seen in his animation (external link) of a bioluminescent millipede walking:
National Geographic: glowing millipede
Note also the palpitating motion of the antennae and that the wavelength of leg movement is
about half the body length in this millipede.





Millipede Taxonomy
Millipede taxonomy is quite complex, but some of the main groups are:
Polydesmids - the largest order with thousands of different species - 'flat-backed' millipedes;
eyeless, e.g. Polydesmus; tergites drawn out into lateral wings called paranota. Platydesmids
are another group of flat-backed millipedes.
Julids - millipedes with cyldindricals egments, e.g. Julus, Cylindroidus.
Sphaerotheriida - giant pill millipedes: roll into a tight ball for defense.
Glomerids - roll into a tight ball for defense, e.g. Glomeris.
Millipede taxonomy is quite complex, but some of the main groups are:
Polydesmids - the largest order with thousands of different species - 'flat-backed' millipedes;
eyeless, e.g. Polydesmus; tergites drawn out into lateral wings called paranota. Platydesmids
are another group of flat-backed millipedes.
Julids - millipedes with cyldindricals egments, e.g. Julus, Cylindroidus.
Sphaerotheriida - giant pill millipedes: roll into a tight ball for defense.
Glomerids - roll into a tight ball for defense, e.g. Glomeris.

Above: transverse section through a millipede (Julus) diplosegment, showing the two pairs of
ventral legs.
ventral legs.
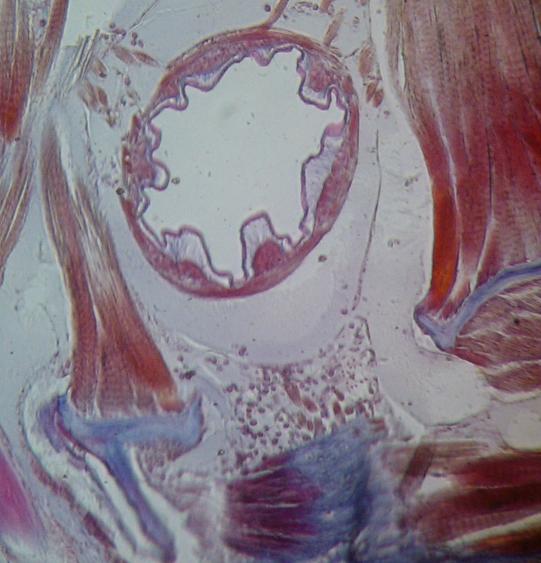
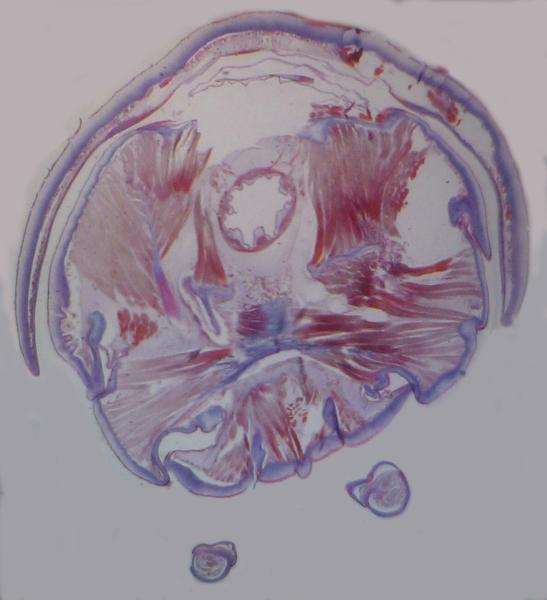
Above: a transverse section through a millipede (Julus). Note the well-developed
muscle fibres attaching to the outer cuticle and to an extensive apodeme system
(ingrowths of the outer cuticle, forming an internal skeleton).
muscle fibres attaching to the outer cuticle and to an extensive apodeme system
(ingrowths of the outer cuticle, forming an internal skeleton).
Above: a close-up view of a cross-section through the millipede gut. Note the
surrounding body muscles and their attachment to the apodemes.
surrounding body muscles and their attachment to the apodemes.
Article updated: 26 Nov 2017