| | | | | | |
| |
| |
| |
| |
| |
| |
| |
| |
| |
| |
| |
| |
| |
| |
| Deep-Sea Fish |
Deep Sea fish come in a variety of fantastic forms with peculiar adaptations to life in the dark depths of Earth's
oceans. We begin our exploration of these bizarre life-forms with the Gulper Eels, such as the Pelican Eel
(Umbrella-mouth eel) Eurypharynx pelacanoides, the only known species in the genus. The term 'Gulper Eel'
also applies to Saccopharynx, another genus of deep-sea fish. Gulpers and Swallowers are a family
(Saccopharyngidae) of ray-finned fish and are not true eels. This mysterious creature is poorly understood.
Below we shall examine some of the remarkable adaptations of the gulpers and other deep-sea fish to life in
the ocean depths. How do these animals find food in the dark ocean depths? How do they find each other for
reproduction? How do they economise in such sparsely populated waters where meals may be few and far
between?
oceans. We begin our exploration of these bizarre life-forms with the Gulper Eels, such as the Pelican Eel
(Umbrella-mouth eel) Eurypharynx pelacanoides, the only known species in the genus. The term 'Gulper Eel'
also applies to Saccopharynx, another genus of deep-sea fish. Gulpers and Swallowers are a family
(Saccopharyngidae) of ray-finned fish and are not true eels. This mysterious creature is poorly understood.
Below we shall examine some of the remarkable adaptations of the gulpers and other deep-sea fish to life in
the ocean depths. How do these animals find food in the dark ocean depths? How do they find each other for
reproduction? How do they economise in such sparsely populated waters where meals may be few and far
between?
Above: A model of the Gulper Eel Eurypharynx.





Eurypharynx pelecanoides reaches a recorded length of 62 cm,
though some gulper eels (Saccopharynx) may be 2 m long. These fish
are found at depths of 500 to 3000 m. Living in the ocean depths, an
ambush predator like Eurpharynx, must make the most of any meal
which comes along and many deep-sea fish have evolved enormous
mouths in order to swallow whatever they can. Despite this,
Eurypharynx has a less-expandable stomach and so generally feeds
on smaller fish, squid, jelly fish, cephalopods, pelagic tunicates,
ribbonworms, seaweed and crustaceans.
though some gulper eels (Saccopharynx) may be 2 m long. These fish
are found at depths of 500 to 3000 m. Living in the ocean depths, an
ambush predator like Eurpharynx, must make the most of any meal
which comes along and many deep-sea fish have evolved enormous
mouths in order to swallow whatever they can. Despite this,
Eurypharynx has a less-expandable stomach and so generally feeds
on smaller fish, squid, jelly fish, cephalopods, pelagic tunicates,
ribbonworms, seaweed and crustaceans.


Feeding
Above: Eurypharynx with its mouth closed and below, with its mouth
open. The enormous mouth cavity can expand to 5 to 11 times the
volume of the rest of the animal! There are small backward-pointing
teeth within the mouth cavity.
Above: Eurypharynx with its mouth closed and below, with its mouth
open. The enormous mouth cavity can expand to 5 to 11 times the
volume of the rest of the animal! There are small backward-pointing
teeth within the mouth cavity.
Bioluminescence
The skin of Eurypharynx is jet black, such that it
would appear practically invisible to any
unfortunate prey or potential predator. However, it
does have photophores (light-producing
organs). The end of the tail is equipped with a
caudal appendage which has translucent walls
and produces a pinkish light, occasionally flashing
red. The lateral line organs, which run in a strip
along each side of the body, have also been
reported to bioluminesce (colour of the light
produced was not specified) though this awaits
confirmation. These strips have been referred to
as 'landing strip lights' since they resemble the
lights on an airfield landing strip. In our model they
have been depicted emitting a pinkish light. If the
caudal appendage is used as a fishing lure to
attract potential prey, as seems likely, then pink
and red seem odd colours to chose as most
deep-sea animals can not see red light. However,
the emitted spectrum might also contain other
wavelengths of light and so appear a different
colour to prey. Alternatively, the light could be
used for other functions, such as communication
or illuminating prey.
The skin of Eurypharynx is jet black, such that it
would appear practically invisible to any
unfortunate prey or potential predator. However, it
does have photophores (light-producing
organs). The end of the tail is equipped with a
caudal appendage which has translucent walls
and produces a pinkish light, occasionally flashing
red. The lateral line organs, which run in a strip
along each side of the body, have also been
reported to bioluminesce (colour of the light
produced was not specified) though this awaits
confirmation. These strips have been referred to
as 'landing strip lights' since they resemble the
lights on an airfield landing strip. In our model they
have been depicted emitting a pinkish light. If the
caudal appendage is used as a fishing lure to
attract potential prey, as seems likely, then pink
and red seem odd colours to chose as most
deep-sea animals can not see red light. However,
the emitted spectrum might also contain other
wavelengths of light and so appear a different
colour to prey. Alternatively, the light could be
used for other functions, such as communication
or illuminating prey.
Above: the caudal appendage has translucent
'windows' through which a pair of black rods are
visible inside. Light is emitted through the windows.
'windows' through which a pair of black rods are
visible inside. Light is emitted through the windows.
It is not known how Eurypharynx feeds, however, it is unlikely that it
swims along with its mouth agape to collect food, since t has only a
narrow pore carrying water out that is exiting the back of the mouth
cavity after flowing across the gills, so it would take too long for
water to exit the mouth cavity and swimming forwards with an open
mouth would be hard work. Furthermore, Eurypharynx has a
narrow body with reduced swimming muscles so is unlikely to be
an energetic swimmer.
It is thought unlikely that Eurypharynx is a 'gape and suck' feeder.
Fish of this type open their mouths suddenly, when prey is in
range, sucking in a stream of water which pulls the prey in with it.
The jaw muscles of Eurypharynx are thought to be too weak to
open the mouth quickly enough.
A more plausible suggestion as to how Eurypharynx feeds is that it
sits in ambush, perhaps in an S-shape hanging vertically ready to
spring into action, and attracts prey with its tail-light. The prey
thinking it has sighted a small bioluminescent animal to eat
approaches and Eurypharynx may then lunge forward with its
mouth agape, engulf the prey and then slowly strain out the water.
Eurypharynx has a low-density and highly buoyant fragile body.
This enables it to hang motionless for long periods of time, waiting
for occasional prey, whilst expending very little energy.
The long jaws and pharynx are adapted to allow the jaws to open
wide. Some of the usual jaw bones are missing and others are
loosely attached to the skull. There are 6 pairs of gill clefts and 5
pairs of complete gills which are supported by the small gill arches
which contain small bones not attached to the skull. The gills are
small, again supporting the view that these fish are highly
economical on their use of energy and exert themselves as little as
possible.
Eurypharynx has two small eyes on the tip of its snout. Despite
being small the eyes are reportedly well-developed and they have
an overlapping binocular field of view which would make them
well-suited to estimate the distance to a prey target prior to striking.
The biomechanics of buoyancy in deep sea fish are summarised in
this pdf: The Biomechanics of Buoyancy in Fish.
swims along with its mouth agape to collect food, since t has only a
narrow pore carrying water out that is exiting the back of the mouth
cavity after flowing across the gills, so it would take too long for
water to exit the mouth cavity and swimming forwards with an open
mouth would be hard work. Furthermore, Eurypharynx has a
narrow body with reduced swimming muscles so is unlikely to be
an energetic swimmer.
It is thought unlikely that Eurypharynx is a 'gape and suck' feeder.
Fish of this type open their mouths suddenly, when prey is in
range, sucking in a stream of water which pulls the prey in with it.
The jaw muscles of Eurypharynx are thought to be too weak to
open the mouth quickly enough.
A more plausible suggestion as to how Eurypharynx feeds is that it
sits in ambush, perhaps in an S-shape hanging vertically ready to
spring into action, and attracts prey with its tail-light. The prey
thinking it has sighted a small bioluminescent animal to eat
approaches and Eurypharynx may then lunge forward with its
mouth agape, engulf the prey and then slowly strain out the water.
Eurypharynx has a low-density and highly buoyant fragile body.
This enables it to hang motionless for long periods of time, waiting
for occasional prey, whilst expending very little energy.
The long jaws and pharynx are adapted to allow the jaws to open
wide. Some of the usual jaw bones are missing and others are
loosely attached to the skull. There are 6 pairs of gill clefts and 5
pairs of complete gills which are supported by the small gill arches
which contain small bones not attached to the skull. The gills are
small, again supporting the view that these fish are highly
economical on their use of energy and exert themselves as little as
possible.
Eurypharynx has two small eyes on the tip of its snout. Despite
being small the eyes are reportedly well-developed and they have
an overlapping binocular field of view which would make them
well-suited to estimate the distance to a prey target prior to striking.
The biomechanics of buoyancy in deep sea fish are summarised in
this pdf: The Biomechanics of Buoyancy in Fish.
Buoyancy
Fish can achieve buoyancy in three principle ways.
The first of these is to rely on lift generated by the
wings acting as hydrofoils (essentially as wings
under water). Sharks use this method which requires
the animal to move constantly or else it will sink. This
is clearly energy costly but can be economical for
fish that are constantly actively hunting for food. The
second strategy is to use a gas-filled swim-bladder
to reduce the rate of sinking, or eliminate it
completely if the animal achieves neutral buoyancy,
i.e., if it has the same mean density as water so that
the upwards buoyancy force cancels the downwards
acting force due to gravity (weight) by 'Archimedes
principle'. The use of gas-filled swim bladders
diminishes with depth, however, since gases are
compressible and their density increases rapidly with
depth; it also takes more energy to fill such a
swim-bladder under high pressure.
Eurypharynx has no swim bladder and with its tiny
pectoral fins it certainly is not built to rely heavily on
hydrodynamic lift. Instead, the Pelican Eel has large
fluid-filled cavities throughout its body, which have
been interpreted as lymph sacs, though there is
some suggestion that they might contain a light
gelatinous material in life. It has also reduced its
denser body components: the bony skeleton is much
reduced. There are no opercular bones (which
support the gill covers or opercula in fishes and the
spine has prominent intervertebral discs made of
persistent notochord in between the biconcave
vertebrae and the body is scaleless. The muscles
also have reduced density, being relatively watery
and gelatinous: as a result the muscles will also be
weaker and the whole animal is designed to hang
motionless in the water column rather than for an
active lifestyle. These adaptations reduce the
density of the animal to, or near to, neutral
buoyancy so it does not need to swim much to stay
afloat.
Fish can achieve buoyancy in three principle ways.
The first of these is to rely on lift generated by the
wings acting as hydrofoils (essentially as wings
under water). Sharks use this method which requires
the animal to move constantly or else it will sink. This
is clearly energy costly but can be economical for
fish that are constantly actively hunting for food. The
second strategy is to use a gas-filled swim-bladder
to reduce the rate of sinking, or eliminate it
completely if the animal achieves neutral buoyancy,
i.e., if it has the same mean density as water so that
the upwards buoyancy force cancels the downwards
acting force due to gravity (weight) by 'Archimedes
principle'. The use of gas-filled swim bladders
diminishes with depth, however, since gases are
compressible and their density increases rapidly with
depth; it also takes more energy to fill such a
swim-bladder under high pressure.
Eurypharynx has no swim bladder and with its tiny
pectoral fins it certainly is not built to rely heavily on
hydrodynamic lift. Instead, the Pelican Eel has large
fluid-filled cavities throughout its body, which have
been interpreted as lymph sacs, though there is
some suggestion that they might contain a light
gelatinous material in life. It has also reduced its
denser body components: the bony skeleton is much
reduced. There are no opercular bones (which
support the gill covers or opercula in fishes and the
spine has prominent intervertebral discs made of
persistent notochord in between the biconcave
vertebrae and the body is scaleless. The muscles
also have reduced density, being relatively watery
and gelatinous: as a result the muscles will also be
weaker and the whole animal is designed to hang
motionless in the water column rather than for an
active lifestyle. These adaptations reduce the
density of the animal to, or near to, neutral
buoyancy so it does not need to swim much to stay
afloat.
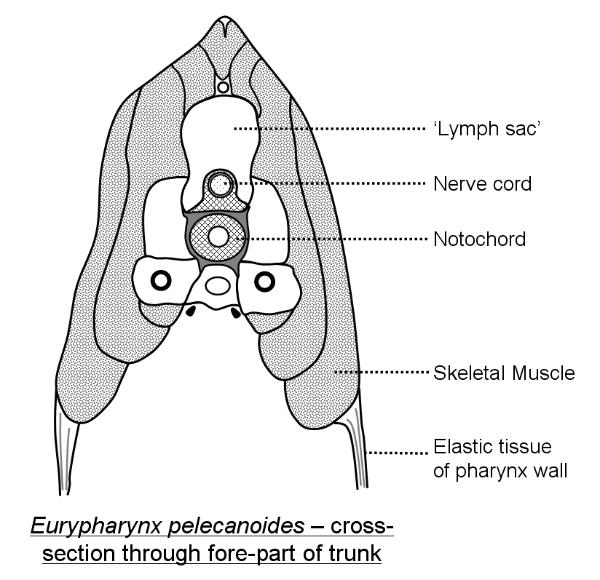
Above: a cross-section through the front end of Eurypharynx: note the extensive 'lymph sacs'
which aid buoyancy.
which aid buoyancy.
The kidneys of those Gulper Eels examined have an
unusual feature: their nephrons lack glomeruli meaning
that there is little filtration of the blood (presumably they
rely more on secretion of materials into the nephrons).
This is thought to be an adaptation to enable the tissues
to retain water, making the animal's tissues watery to
reduce their density.
Reproduction
When ripe, male Pelican Eels develop a large rosette-like
structure inside their nasal cavities, open to the water by
the nostrils or external nares. The nares are at the very
tip of the snout and the development of these internal
rosettes gives the head of the male a notched view when
seen from above. These rosettes are presumably
chemoreceptive olfactory organs and are probably used
to detect pheromones released by the ripe females to
help the male locate them in the dark depths. It is
suspected that Eurypharynx, or at least the female, is
semelparous: reproducing once and then dying.
The eggs hatch into planktonic larvae which live near
the sea's surface, dispersing before descending.
unusual feature: their nephrons lack glomeruli meaning
that there is little filtration of the blood (presumably they
rely more on secretion of materials into the nephrons).
This is thought to be an adaptation to enable the tissues
to retain water, making the animal's tissues watery to
reduce their density.
Reproduction
When ripe, male Pelican Eels develop a large rosette-like
structure inside their nasal cavities, open to the water by
the nostrils or external nares. The nares are at the very
tip of the snout and the development of these internal
rosettes gives the head of the male a notched view when
seen from above. These rosettes are presumably
chemoreceptive olfactory organs and are probably used
to detect pheromones released by the ripe females to
help the male locate them in the dark depths. It is
suspected that Eurypharynx, or at least the female, is
semelparous: reproducing once and then dying.
The eggs hatch into planktonic larvae which live near
the sea's surface, dispersing before descending.
Lateral Line Organs and Sense of Hearing
Fish have an octavolateralis system, the most well-known
component of which is the lateral line of sensors which runs
on either side of the body along its length. The system also
includes, however, similar canals around the head, which
may be very well developed in deep-sea fish and may also
extend along fins and long sensory spines as in certain
anglerfish. Many fish have lateral lines consisting of
fluid-filled canals connected to the surface at intervals by
pores. These serve to detect pressure differences in the sea
water along the length of the animal. Pressure differences
displace fluid within the canals causing minute sensory plates
inside the canals to deflect. This enables fish to sense
disturbances in the water due to movements of nearby fish
as well as certain sounds.
The octavolateralis system of deep-sea fish is often very
different: the sensory plates (forming mechanosensory units
called neuromasts) are not confined inside canals but are
instead borne on the surface of the skin as papillae. This
suggests that instead of detecting pressure waves, they
serve primarily to detect the flow of water particles across the
skin. Sound consists of two components: a near field, which
consists mainly of flowing particles which have been
displaced by the source of the sound. The near field
dominates within about two wavelengths. Beyond this the
more familiar far-field sound, consisting of a density or
pressure wave. Thus it seems that the octavolateralis
systems of deep-sea fishes like Eurypharynx are adapted for
detecting the movements of nearby fish rather than distant
sounds. This suggests they function mainly in detecting prey
within striking range (and predators?).
Near to the skin of a fish there is a boundary layer where
water moving over the skin is changing velocity from the
stationary layer immediately next to the skin to the
faster-flowing mainstream. (This is due to the friction between
the water and the fish's skin). Slower water flows and slower
frequencies of oscillating water flows create thicker boundary
layers. Many deep-sea fish have their octavolateralis sensors
on the tips of minute papillae, column-like or wart-like
structures standing up from the surface of the skin. This
optimises the sensors for the detection of lower frequency
vibrations, presumably corresponding to prey movements.
This must also, however, make it easy for the fish to detect
its own movements. The nervous system of fishes has been
shown to at least partially compensate for this, but hanging
still in the darkness, with the aid of neutral buoyancy must
make it easier to detect other animals nearby.
There is tremendous variation in the construction of the
octavolateralis system in fish and deep-sea fish and the
exact functionality is not well understood, but it would seem
logical that these systems will be tuned to detect predators
and prey (and other members of the same species in
shoaling fish).
Many fish with gas-filled swim-bladders have well-developed
ears which connect to the air-bladders which serve to detect
sounds: as sound waves pass through the fish's body it
causes disturbances in the gas which can be easily detected.
Fish like Eurpharynx which lack swim-bladders may still be
able to hear with their ears, but with greatly reduced
sensitivity. The ears of fish are very different to those of
mammals: they are not connected to the outside by openings
and there is no cochlea. Instead the sacculus which
contains a heavy otolith attached to sensory hairs is the
principle organ of hearing in fish (the role of the
octavolateralis system in hearing is not clear).
Fish have an octavolateralis system, the most well-known
component of which is the lateral line of sensors which runs
on either side of the body along its length. The system also
includes, however, similar canals around the head, which
may be very well developed in deep-sea fish and may also
extend along fins and long sensory spines as in certain
anglerfish. Many fish have lateral lines consisting of
fluid-filled canals connected to the surface at intervals by
pores. These serve to detect pressure differences in the sea
water along the length of the animal. Pressure differences
displace fluid within the canals causing minute sensory plates
inside the canals to deflect. This enables fish to sense
disturbances in the water due to movements of nearby fish
as well as certain sounds.
The octavolateralis system of deep-sea fish is often very
different: the sensory plates (forming mechanosensory units
called neuromasts) are not confined inside canals but are
instead borne on the surface of the skin as papillae. This
suggests that instead of detecting pressure waves, they
serve primarily to detect the flow of water particles across the
skin. Sound consists of two components: a near field, which
consists mainly of flowing particles which have been
displaced by the source of the sound. The near field
dominates within about two wavelengths. Beyond this the
more familiar far-field sound, consisting of a density or
pressure wave. Thus it seems that the octavolateralis
systems of deep-sea fishes like Eurypharynx are adapted for
detecting the movements of nearby fish rather than distant
sounds. This suggests they function mainly in detecting prey
within striking range (and predators?).
Near to the skin of a fish there is a boundary layer where
water moving over the skin is changing velocity from the
stationary layer immediately next to the skin to the
faster-flowing mainstream. (This is due to the friction between
the water and the fish's skin). Slower water flows and slower
frequencies of oscillating water flows create thicker boundary
layers. Many deep-sea fish have their octavolateralis sensors
on the tips of minute papillae, column-like or wart-like
structures standing up from the surface of the skin. This
optimises the sensors for the detection of lower frequency
vibrations, presumably corresponding to prey movements.
This must also, however, make it easy for the fish to detect
its own movements. The nervous system of fishes has been
shown to at least partially compensate for this, but hanging
still in the darkness, with the aid of neutral buoyancy must
make it easier to detect other animals nearby.
There is tremendous variation in the construction of the
octavolateralis system in fish and deep-sea fish and the
exact functionality is not well understood, but it would seem
logical that these systems will be tuned to detect predators
and prey (and other members of the same species in
shoaling fish).
Many fish with gas-filled swim-bladders have well-developed
ears which connect to the air-bladders which serve to detect
sounds: as sound waves pass through the fish's body it
causes disturbances in the gas which can be easily detected.
Fish like Eurpharynx which lack swim-bladders may still be
able to hear with their ears, but with greatly reduced
sensitivity. The ears of fish are very different to those of
mammals: they are not connected to the outside by openings
and there is no cochlea. Instead the sacculus which
contains a heavy otolith attached to sensory hairs is the
principle organ of hearing in fish (the role of the
octavolateralis system in hearing is not clear).
Above: anglerfish like this Bearded Sea Devil
(Linophryne arborifera) (female shown) also share many
of the Gulpers' features: a large mouth and a small body
which is close to neutrally buoyant and a bioluminescent
fishing lure at the end of a rod at the top of the snout to
attract potential prey. The beard that this species has is
also bioluminescent. Many anglerfish are 'gape and suck'
predators. The males of many species are minute
compared to the females and have large olfactory organs.
Having located a female the male attaches, temporarily in
some species, but with permanent fusion of tissues in a
'parasitic' mode of life in others. Attachment is necessary
to trigger ripening of the gonads. Sometimes a female
may have several males attached.
(Linophryne arborifera) (female shown) also share many
of the Gulpers' features: a large mouth and a small body
which is close to neutrally buoyant and a bioluminescent
fishing lure at the end of a rod at the top of the snout to
attract potential prey. The beard that this species has is
also bioluminescent. Many anglerfish are 'gape and suck'
predators. The males of many species are minute
compared to the females and have large olfactory organs.
Having located a female the male attaches, temporarily in
some species, but with permanent fusion of tissues in a
'parasitic' mode of life in others. Attachment is necessary
to trigger ripening of the gonads. Sometimes a female
may have several males attached.
Above: Eurypharynx pelecanoides; below: Saccopharynx. Saccopharynx is another Gulper genus. One of the key
differences is that Saccopharynx also has a highly distensible stomach and so can swallow much larger prey whole. The
caudal appendage on the end of the tail may also be bioluminescent and may be simple or consisting of a series of small
tentacles, depending on species.
differences is that Saccopharynx also has a highly distensible stomach and so can swallow much larger prey whole. The
caudal appendage on the end of the tail may also be bioluminescent and may be simple or consisting of a series of small
tentacles, depending on species.



Bioluminescence in Other Deep Sea Fish
In many fish (as well as in squid and crustaceans)
bioluminescence relies on special light-producing organs
called photophores. Photophores in fish rely on
symbiotic bacteria to produce light. The bacteria only
produce this light when their populations reaches a
critical density and they monitor their own population
density by quorum sensing. Photophores have various
elaborations to modify or enhance their function. Like a
car headlight they may have cup-shaped reflectors to
direct the emitted light in one direction as a focused
beam. Lenses or reflective cylinders may further
collimate the light into a more focused beam. The organ
may be able to shut off the light, either by rotation or by
the action of shutters. Colour filters (which may rely on
pigments or diffractive interference) can change the
colour of the emitted light. Most photophores emit blue
light to maximise the range since blue light travels further
in water which absorbs red light.
Finally, photophores may have wave guides or light
pipes to either spread the light out or carry it to other
parts of the body. These waveguides essentially act as
fibre-optic cables and operate via the same principle of
total internal reflection: the light is transmitted along their
length by reflecting from the inner surfaces of the wave
guide tube. For example, some angler fish may transmit
light from an internal photophore along the stem to the
lure (or accessory structures) used to attract potential
prey.
Most bioluminescence is protective: sudden flashes may
startle potential predators or prevent damage from
accidental collisions. Dragonfishes (e.g. Astronesthes)
produce rapid bursts of defensive flashes of light.
Defensive flashes may illuminate the outline of the
light-emitter, indicating its size to perhaps intimidate.
In many fish (as well as in squid and crustaceans)
bioluminescence relies on special light-producing organs
called photophores. Photophores in fish rely on
symbiotic bacteria to produce light. The bacteria only
produce this light when their populations reaches a
critical density and they monitor their own population
density by quorum sensing. Photophores have various
elaborations to modify or enhance their function. Like a
car headlight they may have cup-shaped reflectors to
direct the emitted light in one direction as a focused
beam. Lenses or reflective cylinders may further
collimate the light into a more focused beam. The organ
may be able to shut off the light, either by rotation or by
the action of shutters. Colour filters (which may rely on
pigments or diffractive interference) can change the
colour of the emitted light. Most photophores emit blue
light to maximise the range since blue light travels further
in water which absorbs red light.
Finally, photophores may have wave guides or light
pipes to either spread the light out or carry it to other
parts of the body. These waveguides essentially act as
fibre-optic cables and operate via the same principle of
total internal reflection: the light is transmitted along their
length by reflecting from the inner surfaces of the wave
guide tube. For example, some angler fish may transmit
light from an internal photophore along the stem to the
lure (or accessory structures) used to attract potential
prey.
Most bioluminescence is protective: sudden flashes may
startle potential predators or prevent damage from
accidental collisions. Dragonfishes (e.g. Astronesthes)
produce rapid bursts of defensive flashes of light.
Defensive flashes may illuminate the outline of the
light-emitter, indicating its size to perhaps intimidate.
Mesopelagic fish live in more shallow waters (down to
about 1000 m) where there is still faint down-welling
scattered sunlight. This poses danger since a dark
silhouette will stand out against the brighter surface when
viewed by a potential predator from beneath.
Hatchetfishes use counterillumination to make
themselves appear invisible from beneath. They have a
series of downwards pointing photophores on the
underside of their bodies. Light is funneled through
silvered tubes and passes through colour filters to match
the colour and spectrum of the down-welling sunlight
exactly. As the intensity of the down-welling light changes
with time of day or depth, the hatchetfish will adjust the
intensity of its emitting light to match. A small pair of
photophores direct light into each eye so that the eyes
can compare the intensity of emitted light to that of the
down-welling light so that any necessary adjustments can
be made. Some squid also use counterillumination.
Many fish probably use bioluminescence to communicate
or signal to one-another. This may be used to attract
mates, to maintain relationships, and to help fish of the
same species to school together. A similar phenomenon
is reported in squid.
Finally, some fish do appear to use bioluminescence to
light-up their path and to illuminate potential prey.
Malocosteus is one such example and this fish has a
red-light emitting photophore beneath each eye. These
fish may emit and be able to see red light which most fish
cannot see, since natural light is green or blue at depth.
Many organisms are also red-coloured since this appears
dark in the sea and will stand out when illuminated by red
light.
about 1000 m) where there is still faint down-welling
scattered sunlight. This poses danger since a dark
silhouette will stand out against the brighter surface when
viewed by a potential predator from beneath.
Hatchetfishes use counterillumination to make
themselves appear invisible from beneath. They have a
series of downwards pointing photophores on the
underside of their bodies. Light is funneled through
silvered tubes and passes through colour filters to match
the colour and spectrum of the down-welling sunlight
exactly. As the intensity of the down-welling light changes
with time of day or depth, the hatchetfish will adjust the
intensity of its emitting light to match. A small pair of
photophores direct light into each eye so that the eyes
can compare the intensity of emitted light to that of the
down-welling light so that any necessary adjustments can
be made. Some squid also use counterillumination.
Many fish probably use bioluminescence to communicate
or signal to one-another. This may be used to attract
mates, to maintain relationships, and to help fish of the
same species to school together. A similar phenomenon
is reported in squid.
Finally, some fish do appear to use bioluminescence to
light-up their path and to illuminate potential prey.
Malocosteus is one such example and this fish has a
red-light emitting photophore beneath each eye. These
fish may emit and be able to see red light which most fish
cannot see, since natural light is green or blue at depth.
Many organisms are also red-coloured since this appears
dark in the sea and will stand out when illuminated by red
light.
Above: an anglerfish with bioluminescent lures. In some anglerfishes a light tube (waveguide)
carries light from the main lure along an accessory tube to be emitted from the tip.
carries light from the main lure along an accessory tube to be emitted from the tip.
Article updated: 10/12/2017