Funaria
(Cord Mosses)

Funaria hygrometrica (Bonfire Moss or Common Cord-moss, Family: Funariaceae, Order: Funariales) is an acrocarpous moss, meaning it develops one fertile 'flower' at the end of each axis. It grows in tufts or clumps on dry heaths and banks, thin soil covering the roots of trees and burnt areas. A variety can also be found on mineral soils, such as cracks in rocks.

The spore capsule is stegocarpous, meaning it splits open (dehisces) along a predetermined line as the operculum (lid) detaches to reveal a peristome consisting of two rings of teeth: the exostome (outer peristome) of 16 teeth, each one opposite one of the 16 teeth (or segments as they are often called in the inner peristome ring) of the endostome (inner peristome). There are no cilia (hairlike appendages) between the segments as there are in many mosses. The outer teeth twist slightly in one direction and arch over such that their tips join to a central sieve-like disc. Thus a cage is formed and, when dry, the slits open wide enough for spores to escape. (Note: stoma means 'opening' or 'mouth' and peristome refers to that which is around the opening, which in this case is the mouth of the capsule through which spores are released).

The calyptra (hood-like remnants of the female reproductive organ or archegonium in which the sporophyte developed, on top of the operculum) is hood-shaped (cucullate) and long rostrate (with a long projecting beak). The calyptra falls as the capsule develops. In at least some varieties, the upper part of the seta (stalk) is hygroscopic and rotates clockwise (when viewed from above) when dry; whilst the lower segment rotates in the opposite sense (counterclockwise when viewed from above) and the middle segment remains untwisted (preventing supercoiling). When moistened, the helical regions uncoil, causing the seta to lengthen. Only mature (non-green) sporophytes respond in this way. When dry, the capsule is angled downwards, ready to drop its spores when the peristome opens, but in moist conditions it straightens up and the peristome closes, conserving spores for more favorable dispersal conditions.

The operculum is caste off when cells in the annulus dry out. The annulus is a ring of cells connecting the operculum to the capsule wall and it is said to be revolute, meaning it uncoils in one continuous strip, detaching the operculum. This leaves the peristome supported on a rim of tissue. The columella is a central cylinder of sterile tissue in the center of the capsule. This breaks down (along with the inner spore sac wall) and the spores around it (in the spore sac) fill the empty space, ready for dispersal. The apophysis (widened part, perhaps more properly called hypophysis) of the capsule has pores called stomata (singular = stoma, since they resemble tiny mouths in shape). When the capsule is developing these pores open and close, opening in daylight and closing at night to allow carbon dioxide to enter for photosynthesis in the light, but conserving water at night. However, evidence suggests that the main function may not be photosynthesis but water evaporation. This evaporation creates a suction pressure which drives the movement of water and nutrients to the rapidly developing capsule, through the conducting strand. As the capsule matures the stomata lose their ability to move and remain open to allow water to evaporate, drying the capsule out as it dehisces (dries and opens) for spore dispersal. The air spaces around the spore sac (supported by trabeculae) assist in drying of the whole structure once it is mature. The stomata of Funaria have a distinct appearance, since the pair of guard cells around the slit-like stoma in most mosses, like a pair of lips, have fused into a single cell, hence we have a slit passing through the surrounding guard cell.

Above: developmental stages of Funaria hygrometrica. A. The main shoot axis ends in a male flower and a lateral branch (usually single) bears the female flower which is beneath the male flower and not yet ripe. B. The male flower is spent and the female flower ripens and its shoot grows to overtop the male flower. Already we see a young sporophyte beginning to develop from a fertiulized egg cell, pushing the end of the flask-shaped archegonium that containe dthe egg, out with it as a beaked hood or calyptra. C. The mature sporophyte, which will dry and open its peristome to shed the spores. The height gained by the seta is enough to clear the most still region of air (the boundary layer) and shed spores into faster moving air. (A and B based on: Boodle, L.A., 1906. The Monoecism of Funaria hygrometrica, Sibth. Annals of Botany, 20(79): 293-299.)
The spore germinates into a tiny filamentous form called the protonema (plural: protonemata) characteristic of mosses. Each protonema puts out between 5 and 27 (or so) gametophores: stalks that develop gametophytes. These begin developing into gametophytes by putting out rhizoids and secondary protonemata from their base. Thus, a single spore can give rise to a clump of mosses. Indeed, Funaria hygrometrica forms such dense patches of moss that insects reportedly find it hard to penetrate the thicket.
During germination, the spore swells with water and greens with chlorophyll. Germination under ideal conditions occurs within one day to a week. Then, a papilla protrudes from the spore surface, until it ruptures the spore coat (exospore) and a protonema grows forth. At first the protonema consists of a single elongated cell, but when its length reaches about 7 or 8 times its width a cross-wall forms, dividing the cell into two. The terminal cell lengthens and divides again, thus a filament or chain of cells is formed. A side branch emerges just behind some a crosswall and reiterates the whole pattern, forming a branched system of filaments.

Funaria hygrometrica is usually autoicous, meaning it has both female and male organs on the same plant. A terminal discoid male 'flower' (male organs or antheridia and surrounding bracts or perichaetial leaves) develops at the end of the main axis. Lateral branches terminate in female 'flowers' (female organs or archegonia and surrounding perichaetial leaves) which are initially shorter than the male flower, when the latter is ripe, but later when the moss enters the female stage these female shoots over-top the exhausted male shoot as seen here. An egg cell in an archegonium of the female flower develops into the sporophyte once it has been fertilized by a motile spermatozoid to form a diploid cell or zygote. Recall that mosses display an alternation of generations, with haploid gametophytes producing the eggs and spermatozoids and diploid spoporophytes forming the haploid spores for dispersal. Occasionally, male and female organs may develop in the same 'flower' (gametoecium) with the archegonia growing singly or in groups of 2-4 surrounded by 15-20 antheridia. In the flowers, sterile hairs called paraphyses pack between the sex organs. In male flowers the paraphyses are straight (filiform) whereas in female flowers they are clavate (club-shaped, with the cells in the clubs well-packed with green chloroplasts). In mixed (synoicous) flowers, filiform paraphyses occur near to the antheridia, clavate paraphyses near the archegonia. (See: Mabel May Brown, The development of the gametophyte and the distribution of sexual characters in Funaria hygrometrica (L.) Scheb.).
The sporophyte is diploid, since it develops from the fertilized zygote by cell division, and consists of the usual foot, seta (stalk) and spore capsule. It obtains nutrients from the parental gametophyte through its foot (it is said to feed parasitically on the gametophyte). The fact that the gametophyte passes through a male phase before the female phase enhances the odds of cross fertilization with another plant. The female branches produce their own anchoring rhizoids and are easily detached to form 'female plants'. Usually each plant produces a single female branch, occasionally more than one, but sometimes the branch is male and itself bears a tertiary female branch.

A gametophyte shoot of phyllidia (leaves) with the gametophyte stem (caulidium) beneath bearing root-like rhizomes for anchorage (one of which is visible in the bottom-left in the above image). The first leaves envelope the apex of the shoot, protecting the growing region. Mosses typically have high regenerative powers. Protonemata (chains of moss cells) can grow from the base of a removed leaf, from stem cells that remain attached.

A single spore can produce a protonema that forms a green felt over the soil surface up to 1 to 6 square centimeters, producing as many as 20 or more gametophytes. Each gametophyte may produce one (occasionally two) sporophytes bearing spore capsules. In this way, Funaria hygrometrica covers a suitable area of soil in a thicket so dense that it is said insects struggle to travel through it.

The spore capsules above are not yet ripe. The operculum has detached, leaving the peristome mounted on the capsule rim, but the teeth have not separated yet (they will when the columella tissue breaks down). The mechanism of the operculum is illustrated below, showing the exostome only:

The exostome teeth, consisting of the cell walls of dead cells, arch upwards and downwards again to contact a central disc (a cribriform or porous disc). In dry conditions, when the spore capsule is angled downwards the peristome opens as the teeth narrow and arch upwards, as shown, widening the slits between them. Each tooth is shaped like a small segment of a helix. It spirals first to the right and upwards.


The arrangement of microscopic fibers in the walls of the peristome teeth causes them to polarize light (Amann, M.J., 1887. On some optical properties of the peristome of mosses. Journal of Natural History, 19(111), pp.248-248).
Below: In damp conditions, when the capsule tends to be upright, the teeth widen and flatten, preventing the spores from being shed in the rain and preventing water entering the capsule and wetting the spores, which may cause them to clump together or decay or germinate inside the capsule.

As already described, the peristome consists of an outer ring (exostome) of 16 outer teeth and an inner ring (endostome) of 16 segments, such that each exostome tooth is opposite an endostome segment. Funaria has a type of peristome called arthrodontous (lit. 'jointed teeth') in which the teeth and segments consist of cell walls from cells that died and ruptured. The peristome forms from 3 layers of cells, the outermost OPL (outer peristomial layer), the middle PPL (primary peristomial layer) and the innermost IPL (inner peristomial layer). A tooth is made up of cell wall material from two cell layers. The exostome teeth consist of the innermost cell wall layer from the OPL and the outermost cell wall layer from the PPL, whilst the endostome segments consist of the innermost wall layer from the PPL and the outermost wall layer from the IPL; the cells themselves having died and ruptured. Thus, each tooth consists of a double membrane (tapering from base to tip) of tapered squares or rectangles (plates of cell wall material) representing the cells from which they were derived. The exact arrangement of the cells that went into making the peristome is of importance in identifying taxonomic groups in mosses.

Above: illustration of peristome segments of Funaria hygrometrica showing the pattern of cell walls that make it up. A: Outermost surface of an exostome tooth, formed from two columns of cells. B: Inner surface of an exostome tooth, derived from a single column of cells. C: Outer face of an endostome segment. D: Inner face of an endostome segment. The fact that the outermost face of the outer ring of teeth presents a double pattern of cell remnants, divided down the middle, makes Funaria hygrometrica a diplolepideous moss (diplolepideous = double scales). In some mosses the outer face is a single column without the vertical dividing midline, these are called haplolepideous (single scales) mosses. The transverse dividing walls are called trabeculae, particularly if they project (like rungs of a ladder).

Above: left, the three layers of cells making up a single developing peristome tooth/segment pair as viewed from beneath (OPL = outer peristomial layer; PPL = principal peristome layer; IPL = inner peristomial layer). Right, a resulting peristome tooth and segment once the cells have died and the cell walls broken apart along predetermined lines.
Peristome formulae. This pattern is different in different groups of mosses and is described by the peristome formula. The peristome formula of Funaria hygrometrica is: 4:2:4. This tells us the number of cells in each layer which make up each of the three wall layers (OPL:PPL:IPL) in one octant (one eight) of the peristome. Since Funaria has 16 teeth (in each ring) this means there are two teeth and two segments per octant, hence 4 OPL cells : 2 PPL cells : 4 IPL cells.
What is the function of the inner peristome?
Not all species of Funaria have the inner peristome. For
example, in Funaria flavicans the inner peristome is reduced,
with very short segments. This moss holds its capsules more-or-less
upright and the seta usually undergoes no or little hygroscopic
movement. The inner peristome acts as a filter, preventing spores from
escaping the capsule too quickly, ensuring dispersal over a period of
time in different wind conditions. In pendant capsules, where gravity
plays a role in spore release, the inner peristome or endostome stops
them falling out all at once. Mosses with more upright capsules
therefore tend to have reduced or absent inner peristomes since gravity
plays less of a role. Many of these mosses may rely on disturbance of
the elastic seta to fling spores out.
Internal structure
A cross-section through the seta (the stalk supporting the
spore capsule) is shown below.
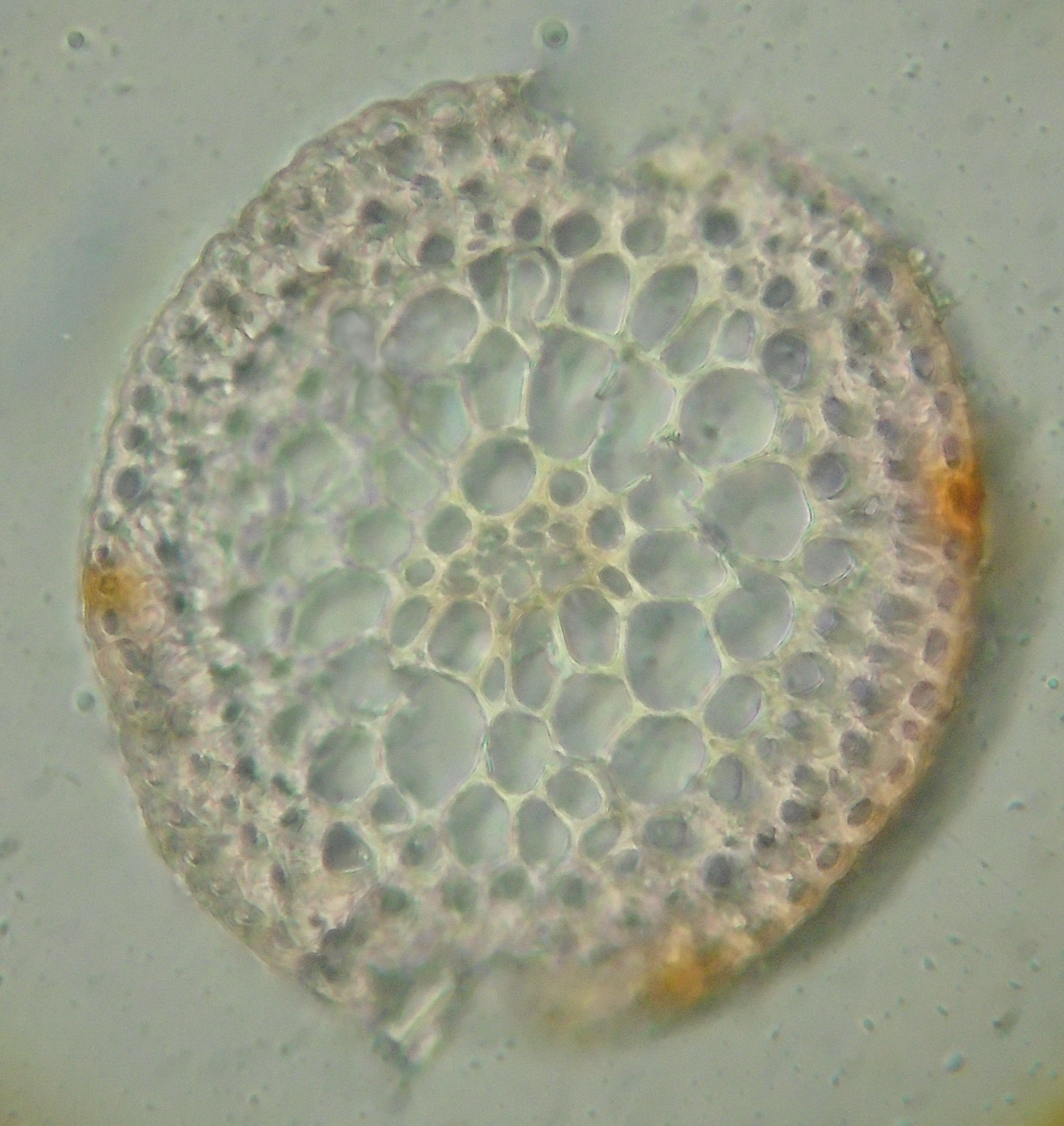
Recall that the seta is part of the sporophyte, the diploid plant that grows 'parasitically' on the haploid gametophyte or leafy moss body. The sporophyte consists of foot, seta and capsule. The seta consists of an outer layer of small, flattened epidermal cells with thick outer walls. The walls of the epidermal cells become impregnated with orange-red pigment when the seta reddens as it matures. Beneath this is the outer cortex, a couple of cell layers consisting of smallish cells with thickened cell walls (these tend to become stereids, cells with very thick cell walls). Inside this are large parenchyma cells. In the center is the central strand of conducting tissue. This consists of a core of thin-walled hydroids, cells that die to leave the empty boxes of their cell walls behind, and these conduct water through their empty lumens, forming pseudo-vessels, not true vessels because there are walls between adjacent cells and water must cross these porous thin walls to pass from cell to cell as it moves up the seta from the gametophyte to the capsule. Surrounding the hydroids there is a sheath of modified cells that have thickened walls, larger diameter than the hydroids and retain their living contents. These are leptoids that convey nutrients, such as sugars. Stereids, hydroids and leptoids may form tissues with some parenchyma cells mixed in and these tissues are referred to as sterom, hydrom and leptom.


British Isles, v.c. 37.
Nigella damascena occurs in the Mediterranean and Near East
Article updated: 28 Dec 2020