| |
|
|
|
|
|
|
|
| |
|
|
|
|
|
|
|
Sponges - Porifera


Above: a Pov-Ray model of a sponge. Sponges belong to the phylum Porifera, which literally many 'many pores' since the surface of a sponge is covered in minute pores that suck in water and nutrients, which the sponge filters before expelling the water from a large opening or osculum. Sponges are usually brightly coloured - red, orange, purple, green and yellow are common sponge colours, though deep sea forms are often dark brown and drab or glassy. The sponge above is definitely an individual - it has a single large opening, or osculum, which carries a jet of water out of the sponge's body, but many sponges form colonies of many individuals fused together with many oscula, in which case it is not very sensible to talk about an individual sponge (although the tissue surrounding each osculum may define an individual). Sponge bodies may be ball, vase, basket, cup or club-shaped like the one above, or they may be flat and encrusting or branching and tree-like. Indeed, the shape of a sponge is an adaptation to its environment - in rough waters, such as along a rocky coastline, sponges will be flat and encrusting, clinging tightly to any nook and cranny they can find in the rock surface, whilst in calm waters they tend to be more upright and present a larger surface to the water to enhance the rate at which they can sieve food from the water. Sponges may be tiny forms, which are easily overlooked, or they may be large enough for a man to stand inside! Sponges are very unusual animals, representing an off-shoot of the animal kingdom that evolved along its own lines, separate from the vast majority of animal types. It is instructive, therefore, to see how the sponge body is put together and how it works!
The simplest type of sponge is the asconoid type. This sponge has many minute pores opening directly into a single central cavity, or atrium (spongocoel), and one or more large oscula (singular: osculum). The sponge sucks water into the atrium through the many tiny incurrent or inhalent pores and pumps it out through the excurrent or exhalent osculum as a forceful jet of water that may travel 10 feet or more from a large sponge. As this water flows through the sponge body, food particles are filtered from it. These particles include mostly bacteria and other microscopic organisms and organic debris. The simplified anatomy of a small asconoid sponge is shown below (most sponges will contain far more cells and many more pores than the simplified model below):

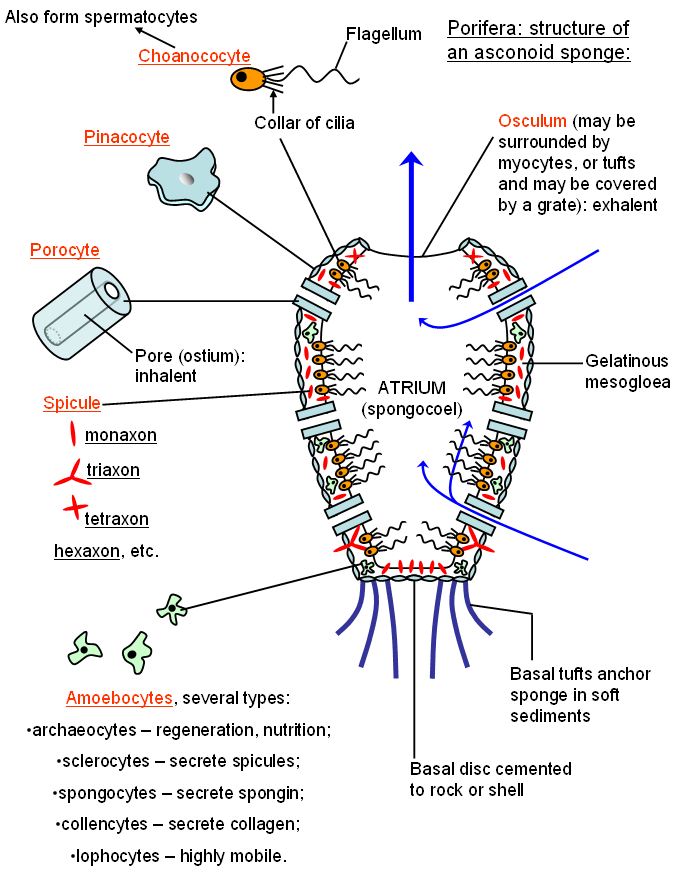
Above: a single module from a type of sponge with the simplest canal architecture: the ascon (asconoid sponge). The asconoid sponge body consists of two layers of cells with a layer of gelatinous material (called mesogloea) sandwiched in-between. The outermost layer consists of flattened paving-slab like cells, called pinacocytes. These cells fit together tightly to form the outer surface or pinacoderm (a type of covering tissue or epidermis). They typically have wavy contours to strengthen the connections between them (by increasing the surface area along which the cells can be fastened together). Inside is a layer of so-called collar cells or choanocytes, each with one flagellum pointing toward the atrial cavity and surrounded by a ring of short appendages called cilia. The flagellum undulates or beats, expelling water out into the atrium and sucking it in through the pores that penetrate both cell layers. Each of these incurrent pores is a channel traveling through a cylindrical cell called a porocyte. Collectively, many thousands or millions of choanocytes can produce a powerful jet of water that leaves the atrium from the large excurrent osculum (or oscula if more than one is present). The choanocytes work very much like a group of organisms called choanoflagellates. Choanoflagellates may live as single cells or in multicellular colonies and are rather like protosponges - sponges lacking the pinacocytes and mesogloea and other packaging cells that make up the sponge body. The structure of a solitary choanoflagellate is shown below:


Above:
the structure and function of a choanoflagellate. These microscopic
creatures beat their flagellum, driving a current of water away from
them and sucking in water from behind which passes between the
collar of microvilli to fill the void left by the water pumped away.
(Note: some refer to the appendages making the collar as cilia,
others as microvilli, but although superficially similar these are
very different organelles). The flagellum has wings or flanges
which help it push against the water more effectively (it must be
remembered that on this microscopic scale water behaves as quite a
thick sticky liquid). As water passes between the collar, potential
food items, like bacteria, are sieved out and ingested by the cell.
This is essentially similar to how a choanocyte in a sponge works
(except that choanocytes lack the flanges on their flagella). Thus,
a sponge's body contains thousands of microscopic pumps that also
filter food from the water. It is thought that choanoflagellates are
related to sponges, and that perhaps sponges evolved from colonies
of these cells.
Another general cell type is typical of sponges - the amoebocyte.
Amoebocytes are amoeboid cells that are free to crawl around (in and
on) the body of the sponge in much the same way as do amoebae. (It
would be interesting to examine the amoeboid locomotion of these
cells in detail to see if it more closely resembles that of amoebae
or of animal cells). Amoebocytes have a variety of functions,
including maintaining the mesogloea (mesenchyme) and removing
foreign organisms and debris from the sponge body. One of the main
functions of the wandering amoebocytes is secretion of the sponge
skeleton.
The mesenchyme
is a transparent gelatinous matrix (mesogloea) containing free
amoebocytes. The mesenchyme may be a collenchyma (meaning it has few
cells and a lot of material between the cells), or a parenchyma
(with a high cell density and little material between the cells as
they are tightly packed together). The amoebocytes are free to
wander about the sponge and fall into two main classes, lobopodous
amoebocytes and collencytes. Lobopodous amoebocytes include
pigmented chromocytes, thesocytes that store food reserves and
scleroblasts that secrete the skeleton. Scleroblasts are further
divided into calcoblasts, silicoblasts and spongioblasts, depending
on the nature of the skeletal material secreted (calcium carbonate,
silicon or spongin protein). Lobopodous amoebocytes have lobopods -
locomotory appendages or psedudopods that are blunt, rounded and
finger-like - the so-called lobopod type of pseudopod. Collencytes
have slender branching pseudopods (called filopods) and may form a
syncytial network (a syncitium is a group of neighbouring cells
whose cell membranes are fused together to form a continuous mass of
cytoplasm. However, distinguishing between a syncitium and a
collection of distinctly 'separate' cells with junctions linking
them together is not easy!).
Archaeocytes are lobopodous amoebocytes. Archaeocytes are possibly
undifferentiated cells and produce the sex cells (as may choanocytes
in some sponges?) i.e. the spermatocytes and oocytes (sperm and egg
cells) and are involved in regeneration since they can give rise to
all other sponge cell types. Indeed, one of the striking
characteristics of sponges is their ability to regenerate from a few
cells - mince
a sponge and each piece can grow back into a new sponge!
Sponge
architectures
The flow of water through a sponge can follow various pathways, giving rise to different canal or aquiferous architectures.
Asconoid sponges (ascons) have the simplest arrangement. The water flows in via the pores, enters the main cavity of the sponge, or spongocoel, which is lined by choanocytes and then exits through the osculum: pores → spongocoel → osculum.
Syconoid
sponges
(sycons) have a more complicated body plan than asconoid sponges.
Syconoids are vase-like with a single, terminal osculum. They have
many finger-like out-pushings which form radial canals continuous
with the central atrium (spongocoel). These external projections
pack the surface and may be free and surrounded by sea-water or they
may be covered by an outer epidermal covering which contains dermal
pores, which open into the channels between the projections, called
incurrent canals, which pass through pores called prosopyles into
the radial canals which are lined by choanocytes. The radial canals
then open into the spongocoel through internal ostia and the water
flows through the osculum to the outside.
Leuconoid
sponges
(leucons) have the most complex structure. There is usually no
central spongocoel, instead the sponge cavity is highly branched and
divided into clusters of small round or oval chambers lined by
choanoflagellates. Mesenchyme fills the spaces around these
chambers. Leuconoid sponges have an indefinite form permeated by a
maze of water channels. Nevertheless, a given unit of water only
flows through one choanocyte chamber as the chambers operate in
parallel rather than in series (there is little point trying to
filter food from water that has already been filtered!). Most
sponges are of the leuconoid type, as this permits the sponge to
develop the most efficient water current and to attain a larger size
(as it filters food from water more efficiently). The leuconoid
architecture is shown in the diagrams below (click the images to
enlarge). This figure was redrawn from Libbie Henrietta Hyman's
classic text: The Invertebrates (Vol 1: Protozoa through
Ctenophora). Note how the choanocytes are restricted to numerous
small chambers formed by the two-fold evagination of the spongocoel.

Above: the principle types of sponge internal architecture. Asconoid sponges are the simplest and smallest and probably the most ancient in design. The whole inner surface of the asconoid is lined by choanocytes (the choanoderm) as indicated in red. The syconoid type represents an array of asconoid types arranged around the cylinder of the sponge (the incurrent pores are not shown, but the red layer is porous in all these sponge types). In type 2 syconoid sponges an additional layer of dermal tissue, perforated by large pores, covers the structure. The leuconoid type is the most advanced and consists of an array of syconoid units arranged around a cylinder. Only the layers shown in red, the choanocyte chambers, bear choanocytes, which line the inside only of each chamber. Syconoid and leuconoid sponges necessarily have a more complex system of pores and channels to convey water through the sponge. The arrangement is such that water only flows through one choanocyte chamber on its passage through the sponge, otherwise energy would be wasted in filtering water that has already been filtered. The more complex syconoid and leuconoid architectures are more efficient and so are able to achieve greater size - an asconoid becomes increasingly less efficient at larger size. Asconoid sponges are small, and rarely exceed 10 cm in height, but the more complex and efficient leuconoid sponges may reach two meters in height. The architectures shown above are all open architectures, in which water exits through a large central spongocoel cavity. Many sponges have solid architectures, in which a series of channels coalesce to expel water through one or more oscula and the large central spongocoel is essentially absent. Leuconoid sponges are the most common and their architecture may be complicated, such that the sponge's tissue becomes permeated by a complex maze of water channels.

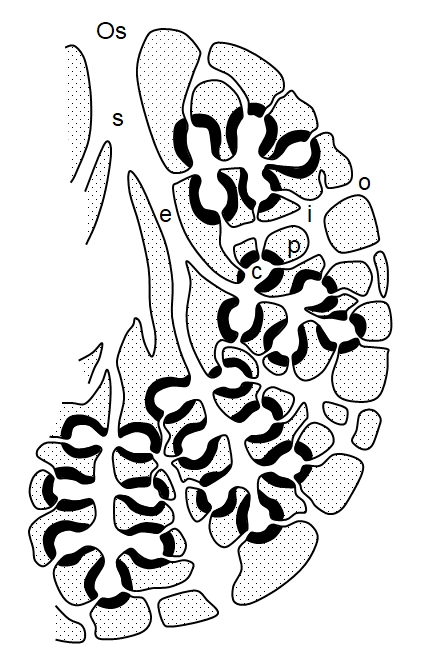

Above: the architecture of a leuconoid sponge (after Hyman, 1940). In many sponges there is variable architecture, but the general pattern seen here involves water flow through the sponge as follows: ostia (o, dermal pores) → incurrent canal (i) → prosopyles (p) → choanocyte (flagellated) chambers (c) → excurrent canal (e) → spogocoel (s) → osculum (Os). This type of arrangement in which the choanocyte chambers connect directly to the excurrent canals by wide apertures is termed eurypylous.
Further modifications may occur, a narrow canal (aphodus, plural: aphodi) may connect each chamber to the excurrent canal, an arrangement termed aphodal: ostia → incurrent canal → prosopyles → choanocyte (flagellated) chambers → aphodus → excurrent canal → spongocoel → osculum. Finally the prosopyle may extend into a narrow canal, such that each chamber has a narrow inlet canal (prosodus, plural: prosodi) and a narrow outlet canal (aphodus) - an arrangement called diplodal: ostia → incurrent canal → prosodus → choanocyte (flagellated) chambers → aphodus → excurrent canal → spongocoel → osculum. In reality an individual sponge may show mixed or intermediate stages of canal architectures. Fort example, a single specimen of Scypha ciliata can show ascon, sycon and leucon types.
Sponges are generally modular organisms, made up of repeating aquiferous modules, and several oscula may be present, one per module. In some sponges, the true oscula themselves open into an atrium which opens to the outside water via a pseduosculum. Conversely, some sponges flatten out with a wide crater-like osculum and may become so wide and flat, with such a low oscular rim, that the osculum essentially disappears and excurrent canals then open directly to the outside. The ostia may aggregate into pore sieves in some sponges.
There are issues over naming the various parts of the aquiferous system of sponges. In particular the ostia are sometimes referred to as dermal pores, but these should not be confused with the pores of the ascon sponge which consist of porocytes.
There are important mechanical factors to consider when understanding the sponge aquiferous system. For example, in a leuconoid sponge, as water flows down through the incurrent canals it is siphoned off into adjacent choanocyte chambers and to maintain filtration pressure along its length, the cross-sectional area of the incurrent canals decreases, maintaining pressure as fluid volume contained within the incurrent system drops. Conversely, as more and more water is pumped into the excurrent canals, they increase their cross-sectional area so as not to generate increasing back pressure which will oppose further filtration (Larsen and Riisg�rd, 1993).
Why the complexity?
This can best be understood in terms of the diameter of supply
(DoS). The sponge must expel water far enough from itself such
that it takes in very little of the water it exhaled, since exhausted
water is depleted of oxygen and food and may contain toxic waste
products. If the exhalent jet is fast enough then it will jet high
enough to avoid all but trace amounts of such recirculation: the sponge
has achieved a sufficient DoS (see diagram below). In a fast current, a
flat encrusting sponge may have an infinite DoS, since the current will
bring in a constant supply of fresh water whilst expelling waste water
far downstream.

After Fry, W.G. , 1979. Taxonomy, the individual and the sponge. In
Biology and Systematics of Colonial Organisms, Larwood, C and Rosen,
B.R. (eds). Academic Press.
To increase DoS a sponge can raise the osculum by placing it at the end of a tall chimney or increase pumping pressure to increase the velocity of the exhalent jet. The pumping pressure of a choanocyte chamber is inversely proportional to its volume: a large single chamber (as in asconids) will generate a lower pressure than a series of smaller chambers arranged in parallel (as in a leuconoid sponge). Thus, the more complex architectures allow sponges to attain a greater size without compromising the DoS. Additionally, splitting the incurrent water and passing it through several smaller filtration chambers, rather than one large chamber, undoubtedly increases filtration efficiency in large sponges.
Most, if not all, sponges can regulate the diameter of the osculum and also regulate water pumping by contracting the sponge body to reduce the volume of the choanocyte chambers. Indeed, many sponges periodically contract so as to completely shut down pumping for transient intervals. Some sponges narrow the osculum aperture by general body contraction, others by contracting the pinacoderm around the osculum but some have specialised muscular sphincters. Generally, as a sponge increases pumping rate it increases opening of the osculum, so maintaining a constant exhalent jet velocity. The oscula may close in still water to prevent the sponge taking in waste water when the DoS is insufficient.
In still water some sponges will grow longer chimneys bearing the oscula at their summits, increasing DoS. In some sponges, several oscula may be grouped together to generate a combined jet which can push higher into the surrounding water (by reducing frictional losses in the compound jet) and oscula may even fuse into single chimneys. Fan-shaped sponges may further solve the problem by orienting into prevailing currents with the ostia on one surface (facing the incoming flow) and oscula on the other surface. Thus, the concept of DoS can go along way to making sense of the complexity of sponge architecture and shape.
The
Sponge Skeleton
The
sponge skeleton deserves special attention. The mesenchyme secretes
and contains the skeleton. The skeleton consists of spicules and/or
spongin fibres. The spicules may be principally calcium carbonate in
calcareous sponges and are made of silica in siliceous sponges;
spongin is a protein. The spicules
(sclerites) are
tiny (mostly microscopic) crystalline bodies and each is a spine or
a number of spines radiating from a point. Each spicule consists of
an organic axis surrounded by calcium carbonate or hydrated silica.
Megascleres are the larger spicules
that from the main supporting framework. Microscleres are smaller flesh spicules
strewn throughout the mesenchyme. However, such a size distinction
does not hold for calcareous sponges and some other groups.
Spicules are classified according to the number of spines or axes as
follows (this topic gets very technical and has been much
simplified!):
1. Monaxon
spicules
have a single axis, straight or curved. There are many types
according to their shape and how they form. They may be lance-like,
C-shaped, bow-shaped, thorny or knobbly, rod-shaped, twisted spirals
or spiny. Some have a pointed end projecting to the exterior of the
sponge, making its surface rough and spiny, presumably for
protection.
2. Tetraxons (tetractines, quadriradiates) have four rays radiating from a common point. These rays are not in the same planes and may resemble jacks, though some of the rays may be reduced, absent or modified into discs.
3. Triaxons (hexactinal spicules) have three axes crossing at right-angles to give six rays, some of which may be lost or reduced or branched or curved and may have spines or knobs, etc. These spicules occur only in the class Hexactinellida.
4. Polyaxons have several equal rays that radiate from a central point and may be star-shaped or resemble spiny spheres.
5. Spheres form from concentric growth around the centre.
6.
The desma is a megasclere formed
from a minute monaxon, triradiate or tetraxon spicule, forming a
central structure upon which layers of silica are deposited. These
deposits develop branches and tubercles. Desmas are usually united
into a network to form a net-like skeleton (lithistid).
Spongin is a protein that forms a
branching network. In some sponges, spongin often binds the
siliceous spicules together. In the keratosan sponges the skeleton
consists entirely of spongin (and embedded foreign particles).
Spicules are secreted by scleroblasts, which are a sub-class
of amoebocyte. Silicoblasts are a type of scleroblast that secrete
siliceous spicules in the siliceous sponges. Spongin is secreted by
spongioblasts. The various spicule types (of which there are many
sub-types I have not mentioned here) have different skeletal roles
within the sponge body and the types present also depends upon
species. I think that you get the point that the sponge skeleton is
actually rather complex! The spicules make the tissue of many
sponges very hard, prickly or stony and quite difficult to cut with
a knife. Traditional bath sponges (not the artificial type!) have
spongin skeletons and no mineral spicules, which gives them a
springy and spongy texture.
The
main function of the sponge skeleton is to support the aquiferous
system and keep the water canals and chambers open. Individual
choanocyte chambers may be supported by fibers connecting them to
the spicules. The skeleton also functions to protect the sponge:
they often give the sponge a tough texture and spiculues may guard
openings such as the oscula and spicules may project out from the
sponge surface to make them spiny or prickly. The spicules may be
loosely embedded in the fibrous tissues of the sponge or cemented
together. The exact arrangement and the degree to which the spicules
interlock or work together are not always obvious. In forms that
dwell on soft bottoms, long bundles of spicules may protrude from
the basal end as rooting fibers. other forms may attach to hard
surfaces by means of secretion, sometimes spongin, whilst in others
the main body is buried in the sediment with chimneys to carry the
openings clear.
Glass
Sponges (Hexactinellids)
Glass
sponges have a skeleton consisting of a lattice of glass-like
siliceous fibres. These strange sponges are deep-sea sponges,
occurring at 100 m to 5000 m depth. Most are 10-30 cm tall, but some
are over 1 metre. They are pale in colour and may have projecting
spicules (which may be up to 30 cm long) which may give them a
'glass wool' like covering. In at least one form it would appear
that the projecting glass fibres (the spicules) act as fibre-optic
wires, transmitting light from a bioluminescent crustacean
resident that lives inside the spongocoel, which must create a
spectacular light display. It is quite possible that many glass
sponges are bioluminescent. It has also been suggested that the
glass spicules may be part of a light-sensing system, whether for
releasing or receiving light, they are very good at transmitting
light. These mysterious sponges are not very well studies and many
glass-sponge wonders surely await discovery. One well known example
is the Venus's flower basket - the beautiful glassy skeletons of the
sponge Euplectella.
The internal structure of glass sponge tissues is very different from
that of other sponges. They are covered by a dermal membrane,
comprising a syncytium with underlying membrane of spongin (a
type of collagen) and have a very open body plan with large open
spaces, rather like a honeycomb. (A syncytium is a large multinucleate
structure). The 'archaeocytes' are generally connected to one another
by slender cytoplasmic bridges, forming a meshlike or trabecular
syncytium that constitutes the bulk of the sponge body. This
syncytium is continuous with choanablasts, forming a choanosyncytium.
Each choanoblast gives off one or more stalk-like stolons that connect
to collar bearing units, resembling choanocytes but lacking nuclei
(there cytoplasm is continuous with that of the choanoblasts). Stolons
may also connect these collar units to one another. This arrangement
is reminiscent of certain colonial choanoflagellates, in which the
cells are connected by stalk-like or root-like stolons. The only cells
that are routinely seen separated from the syncytium are the scleroblasts,
which secrete many of the spicules. All the other cells appear to be
connected to one another by cytoplasmic bridges, though they may have
rounded bodies, making them resemble distinct cells. It has been
argued that glass sponges are so different to all other sponges that
they should be in a separate phylum. However, their strange internal
architecture stems from the tendency of their cells not to separate
fully at cell division, but to remain in contact with one another by
cytoplasmic bridges (which may be occluded by pore plates that allow
certain materials to pass from 'cell' to 'cell' directly as the
cytoplasm is continuous across the pores) with a single continuous cytoplasmic membrane surrounding the whole
structure. Considering that some choanoflagellates are solitary, whilst
others form colonies of connected individuals, I don't personally think
that this difference is great enough to warrant placing glass sponges in
a separate phylum.

A glass sponge.
Demosponges
Demosponges
either have no spicules at all or they have the siliceous type. All
are of the leuconoid type. There are at least three types, including
the horny sponges (keratosans) which possess spongin skeletons but
have no spicules (unless these are made of spongin), though they may
incorporate rock grains and other materials into their bodies. The
spongin forms a lattice or tree-like branching network throughout the
keratosan sponge body. Keratosans are usually black in colour and have
a smooth or warty leathery texture. They live attached to hard
materials, glued in place by spongin secretion.
Calcareous
sponges
These
have calcareous spicules and include all the asconoid types, some
syconoids and some leuconoids. The calcium carbonate spicules are
usually separate but are sometimes fused into a network or are
enclosed in calcareous cement. Projecting spicules often give these
sponges a bristly texture.



A barrel sponge releasing sperm.
The
shapes of sponges
Sponges
appear in a huge variety of forms: spherical, conical, club-shaped,
vase-like, tubular, goblet or cup-like, encrusting sheets, upright
sheets, plate-like, fans and treelike forms, etc. Sometimes
individuals of the same species, but growing in different locations,
may show very different shapes, although each species tends to adopt
a particular range of shapes. There are several physical reasons why
sponges are the shapes they are:
1. Treelike forms branch in such a way as to maximise the filtration
of water. Just as an oak tree has to maximise the light it
intercepts and the carbon dioxide it removes from the atmosphere, so
a sponge has to maximise the efficiency with it extracts food from
water by filtration. If two branches of an oak tree are too close
together, then their leaves may overlap and the shaded leaves may
receive insufficient light for photosynthesis and they are a waste
to the plant.
Additionally, the leaves of one branch absorb carbon dioxide from
the atmosphere and create a zone of air around the branch depleted
in carbon dioxide which is only slowly replaced by diffusion in slow
air, but is replaced more quickly in windy conditions. With two
branches too close together the branches will compete for carbon
dioxide and most likely there will not be enough to go around. This
would be the case if the crown was a solid ball with no branches -
all parts of the crown would be competing for insufficient resources
and this would be wasteful. To overcome this problem, tress branch
according to one of several optimum branching patterns (the exact
solution used depends upon species, which is one reason why
different species have different branching patterns) which spreads
the branches apart so that they do not compete too much with one
another and the tree can maximise its exploitation of light and
carbon dioxide without wasting resources (like wood).
It is possible to use a computer to solve the mathematical problem
of supply and predict the optimum branching patterns. This has been
done for sponges (using the diffusion equation to model the
diffusion of food particles into the volume of water around the
sponge that the sponge can exploit by sucking this water through its
body). The assumption is that sponge tissue will not grow into
regions of water where it finds insufficient food (such as too close
to other sponge tissue which have already removed the food from the
water). These models correctly predict many sponge-like shapes,
including the branching tree-like forms. One also has to consider
whether or not the sponge lives in the open where there are strong
currents to bring in fresh food rapidly, or whether it lives in a
sheltered region of more stagnant water.
Another factor which effects sponge shape is the diameter of
supply. This diameter is the furthest that a sponge can eject
water from itself before some of that water recirculates and gets
taken up again by the sponge, assuming stagnant conditions. Ideally,
the sponge would take up fresh water without mixing in any of the
water it has already filtered - filtered water has had the food and
much of the oxygen removed from it by the sponge and is now of
little use
and hence the further the sponge can eject it, the better, as then
it is less likely to return or is thoroughly mixed and diluted with
fresh water. In strong currents, this is not a problem, the currents
carry away the waste water and bring in fresh water. Such a sponge
may be an encrusting film that coats the surface of a rock and puts
up very short chimneys, each with an osculum on top. In more
stagnant water, however, the sponge may have taller chimneys
through which to eject its waste, which may help the ejected water
reach the currents higher above the surface (a boundary layer of
stationary or slow-moving fluid always covers the surface of
objects, and the depth of this boundary layer increases in more
stagnant conditions and reduces in strong currents; above the
boundary layer is the turbulent layer where currents mix the water).
Factories use the same principle - taller chimneys carry the waste
higher into the winds where they carried further away.
A third factor determining sponge shape is mechanical. A tall
fan-shaped sponge may be swept away in strong currents, whereas a
slender conical sponge or a low-lying encrusting sponge stuck to the
rocks over a large surface, are more likely to survive. On the other
hand, in milder currents, the flat encrusting form may be trapped
inside the boundary layer, away from the currents higher up that
bring in fresh water, oxygen and food.
Different species are best adapted to different conditions, however,
the fact that sponges can alter their shape to some degree proves
that sponges are sensitive creatures - able to respond to their
environment and grow to a shape that best suites the environment.
For example, the sponge Hymeniacidon
perleve
grows as a thin film that encrusts the surface and puts up a number
of very short chimneys, but sometimes it can grow tall chimneys,
which may branch or
fuse into a single large cluster of chimneys (for greater strength
allowing them to grow taller). Presumably, these different forms are
adaptations to the local environment. Other sponges may grow on
elongated stalks, enabling them to reach the currents in the
turbulent zone and perhaps to avoid sucking up silt from the
sediment, which may clog their pores.

Above: a model of a goblet sponge - several species adopt a cup-like form, complete with stalk and attachment disc. This is one example of an open sponge architecture, in which the outflowing water enters a common spongocoel chamber before being expelled through the osculum. Other sponges have a solid architecture, in which canals permeate the whole body and coalesce to open directly into one or more oscula. Click the image to enlarge.
Regeneration
Sponges have
very high regenerative powers. Any piece can regenerate into a whole
sponge, but the process is slow, requiring months or years for the
new sponge to reach full size. If a sponge is broken into cells and
cell-clumps, then the amoebocytes aggregate to form a reunition
mass. Some reunition masses contain collar cells without collars and
various types of amoebocyte. Some of the amoebocytes form an
epidermis and a whole sponge is reformed. Reunition masses composed
entirely of choanocytes cannot reform a sponge. Cells from different
species, mixed together, may temporarily form a reunition mass
before separating.
In adverse conditions, many marine sponges and freshwater sponges,
collapse and disintegrate to leave a reduction body remnant
comprised of a covering epidermis and an internal amoebocyte mass
with partially de-differentiated choanocytes. This will grow into a
sponge when and if favourable conditions return.
Infaunal
Sponges
A
peculiar type of sponge that is little known and little studies is
the infaunal sponge that lives mostly buried in soft sediment at the
bottom of the sea or below reef slopes on the continental margin.
For example, the model sponge shown below (click thumbnails to
enlarge) is similar to species of Oceanapia consists of a dark brown
spheroidal central bulb, some 6 cm by 4 cm, buried beneath 5-10 cm
of sediment. From this bulb extend 4-9 tubes or siphons, 6-28 cm in
length and about 1 cm in diameter and whose white tips extend 3-8 cm
above the sediment surface. In Oceanapia
peltata,
found off the Colombian coast, these siphons may bear a number of
stacked horizontal discs partially enclosing them (not shown). Also
from the bulb extend a number of tubular 'roots' which penetrate
more or less vertically into the sediment to a distance of several
cm. The description given is intended to assist your imagination,
but infaunal sponges come in a diverse variety of shapes and forms.
Experiments with Oceanapia have shown that the tips
of the siphons are porous and draw water in to a series of channels
that permeate the non-porous bulb. Food particles are filtered from
the water and then the water is expelled into the sediment by the
root-like excurrent tubes (which bear pores of an uncertain nature
as it is hard to collect the specimens without damaging the roots).
In life, these sponges are firm and somewhat elastic, but they
become brittle when dry. The secret nature of
these sponges and the technical difficulties involved in collecting
intact specimens has hampered the study of these sponges. Remember,
that despite their appearance, texture and limited movements,
sponges are animals!

Above:
Some deep-sea sponges are carnivorous! This is Cladorhiza, a carnivorous genus of
sponges. This is a (monaxonid) demosoponge with monaxon
megascleres. Further examples of these bizarre carnivorous sponges
include the filamentous Asbestopluma (resembling a mass of
crystalline filaments), Chondrocladia
lampadiglobus (with a central stem bearing stalked
globules) and Chondrocladia lyra (resembling an 'alien'
harp). These sponges have surfaces which are sticky, sometimes by
means of a layer of tiny protruding hook-like spicules, or perhaps
by glue-like secretions. Small animals, such as shrimp, get
accidentally trapped when they contact the sponge and the nearby
cells of the sponge then move to enclose the prey in a digestive
cavity! This Cladorhiza has a long stem which embeds in the
bottom ooze. The umbrella-like spines possibly serve to both prevent
the sponge sinking too deep into the ooze and to catch prey.

Above: a model of an infaunal sponge such as Oceanapia. These sponges vary considerably in form between species and many undoubtedly remain unknown to science. Siphons emerge above the sediment which buries the bulb-like body and root-like appendages extend into the substrate.
Below: the sponge as seen in its natural habitat - with only the
porous tips of the siphons visible above the sediment.

An interesting paper by Cerrano et al. (2007) discusses the inclusion of foreign objects into the bodies of certain sponges, such as the infaunal sponge Oceanapia fistulosa. Some sponges deliberately incorporate particles of detritus into their own bodies. These particles may come from the 'snow' of detritus that constantly falls to the sea bed or from substrate particles. Infaunal sponges, and certain sponges or fragments of sponges that are normally attached to solid substrates but become dislodged must ensure that they maintain the correct orientation to prevent sediment from clogging their aquiferous system. They may do this by incorporating particles of sediment into their basal portions, acting as 'ballast' and stabilise the sponge body in the correct orientation.The anchoring basal strands of an infaunal sponge like Oceanapia are particularly active in taking in the larger foreign particles (those above about 2 mm are preferred) and may reach a length of 15-20 cm (the globular sponge body may reach 5-15 cm in diameter) and 1 cm in diameter, but these tend to be narrower, longer and more numerous in finer-grained sediment.

This type of particle inclusion seems non-specific, but other parts of sponges may incorporate selective particles, selected on the basis of size and/or mineral composition. When debris falls upon a sponge, the pinacocytes may remove it by wave-like movements of their cell membranes or they may engulf the particles and sort them. Acceptable particles may then be incorporated into the underlying tissues to aid in skeletal support or to give the sponge body toughness. Some species produce no spicules of their own and incorporate foreign spicules and other particles into their spongin skeleton or other matrix should they lack spongin also. Some sponges incorporate silica in the form of quartz, which seems detrimental to some species but beneficial to others. Indeed, some sponges dissolve internalised quartz as a source of silica and in Chondrosia silica switches on the genes for collagen synthesis.
Individuality
in sponges?
Many
sponges are colonial - many tubes, each bearing an osculum vent
may be fused together at their bases to form a single mass. It may
be that each unit, comprising a single osculum and surrounding
tissue is one individual and that one sponge produced new
individuals that failed to separate completely. On the other hand,
it is known that some sponges if placed next to one another will
fuse together to form a single individual, whilst others may fuse
temporarily before rejecting one another, in which case a space
(zone of non-coalescence) appears between them. It turns out that
usually only sponges of the same species and the same strain will
fuse together. A sponge will fuse with pieces of itself - if a
piece of tissue is removed and then placed near to the source
sponge, or inside a hole cut into the sponge (an autograft), then
the fragment will fuse with its parent body. If a graft (say a
cube of tissue) from another sponge is placed inside a hole cut
into the recipient sponge, then only if the graft was from another
sponge of the same strain will it fuse with the recipient. If the
graft is from a different strain, even one of the same species,
then rejection will occur - the graft will not fuse and may become
black and necrotic and shrink away as tissue from the recipient
grows into the wound to replace the rejected graft. Other times
the graft will grow and enlarge at the expense of the recipient
sponge. Biochemicals, called sponge factors, are known to be
secreted by sponge tissues when a graft is introduced, if the
strains are incompatible, then these factors will reduce the
adhesiveness of the cells to one another and the graft will be
unable to stick and fuse to the recipient. Whether we define an
individual sponge as a physically separate sponge (even if one
grew from a fragment of the other) or whether we define the
compatible strain as the individual is a matter of opinion.

One sponge or three? Click image to enlarge.
The
sponge spicule skeleton is a truly remarkable structure that ought
to be the envy of engineers. It is interesting to look at sections
of sponges and see how the various spicules fit together like
mechano into diverse large-scale meshworks designed to take the
weight of the animal, give its tissues hardness and protect it
against predators, and all with minimum material costs and
light-weight construction.
Megascleres
are larger and often mesh together to form the bulk skeleton; microscleres are smaller and
generally free in the tissues, e.g. microcalthrops - a small
calthrops. Triaenes are tetraxons with one long ray, called a
rhsbdome and 3 smaller clads that form the cap-like cladome.
Typical examples of spicule (sclerite) types are illustrated below with 3D computer-generated models:

Two-rayed diaxon.

One-rayed monaxon.

Triaene tetraxon

Amphidisk

Six-rayed triaxon (6 points and 3 axes)



Triaxon (3 rays/axes)

Pentaradiate (5 points and 3 axes)

Tetraxon (4 axes/rays)

Tetraxon (calthrops - equal rays)

An open cubical cage. Several such subunits may be joined into a larger cube unit.

Above and below: spicules.



Above: gemmules.

Above: spongin fibers.
References
Cerrano, C., Calcinai, B., Di Camillo, C.G., Valisano, L. and
Bavestrello, G. 2007. How and why do sponges incorporate foreign
material? Strategies in porifera. Porifera research: Biodiversity,
Innovation and Sustainability: 239-246.
Fry, W.G. 1979. Taxonomy, the individual and the sponge, in Biology and Systematics of Colonial Organisms, Larwood, G. and Rosen, B.R. (eds.). Academic Press.
Hyman, L.H. 1940. the invertebrates: Protozoa through Ctenophora. McGraw-Hill Book Company, Inc. New York and London.
Larsen, P.S. and Riisgård, H.U. 1993. The sponge pump. J. Theor.
Biol. 168: 53-63.
Article
updated: 23/4/2014, 17/11/2019
Article last updated: 9/5/2020