
| Gastropod
Molluscs |
| |
|
|
|
|
|
Above: a
model of a gastropod of the dog whelk type. Notice the muscular
foot, the distinct
head with a pair of eyes, cephalic tentacles and a proboscis. To the animal's right is the large
siphon, whilst to the left is the penis tucked away inside the shell. Notice the operculum or
'shield' on the back of the foot.
Sea Snails
The gastropods are molluscs and so are included in the same phylum as cephalopods and
bivalves. The name 'gastropod' literally means 'stomach-foot' in reference to the way snails
and slugs appear to crawl on their bellies. In reality the stomach and other parts of the gut are
contained in the dorsal hump (beneath the protective shell in snails). Most snails belong to a
group of the gastropods referred to as prosobranchs. Prosobranchs are a diverse group
with multiple ancestors, and so gastropod classification has been overhauled in recent years
to reflect evolutionary relationships. Though useful to studies of evolution, evolutionary
classifications are of limited field use and we shall stick only to descriptive terms in this article,
avoiding complex issues of 'family trees'.
The prosobranchs are 'fore-gilled' snails. What does this mean? Well, in ancestral forms the
anus and pair of excretory pores (nephridiopores) opened at the rear in the mantle cavity.
The mantle cavity is an external space just underneath the rim of the shell, lined of course
by the epithelium (or 'skin') of the snail. A pair of external gills (borne of the surface of the
snail's body) also opened into the rear of this mantle cavity. Thus the gills are in a protected
cavity beneath the shell but open to sea water in the mantle cavity. During the course of
evolution, the gastropod snails underwent torsion. In the developing snail the hump, with its
shell and internal organs, twists through 180 degrees, so that the anus, excretory pores and
gills now sit above the head in the anterior mantle cavity.
The Gastropod Shell
There are some 85 000 described species of snail, though this probably represents a small
fraction. Most are marine, some are freshwater and some are terrestrial. The shell consists of
a cone wound into a spiral about a central axis or columella. Each turn of the spiral is a
whorl and whorls are distinguishable in most cases by external grooves or lines in the shell
called sutures, which may be simple curves or wavy boundaries. The basal whorl is the
largest and called the body whorl and its boundary is the open aperture through which the
foot and head can be protruded or retracted. This aperture defines the anterior or head-end
of the shell, whilst the apex of the shell is posterior. Delimiting the aperture is an outer lip of
shell opposite the inner lip, the latter is continuous with the columella. The smaller (and
older) whorls of the shell form the spire, which decreases in size towards the pointed apex.
The shell surface may be smooth or sculptured with ridges, grooves, ribs, tubercles or spines.
Many marine forms are brightly coloured with spiral bands, blotches, stripes or dots. These
materials may be deposited waste products, or they may have a camouflage function and
include melanins, pyrroles and porphyrins.
The shell may bear a siphonal canal at the front through which the siphon (if present) can
irrigate the mantle cavity with sea water to facilitate gas exchange at the gills and the ejection
of waste. The siphonal canal is an extension of the body whorl and aperture. Opposite there
may be a similar and smaller posterior canal.
The shell may be long and slender with many whorls or short and plump with a reduced spire.
The spire may also be incorporated into the body whorl and so not readily apparent. All the
whorls may lie in one plane (a planospiral shell) or it may be helical. Limpets have conical
shells with no spire and an aperture that occupies the whole width of the shell. In cowries, the
aperture is reduced to an elongated slit.
The mantle, which is the soft tissue lining the mantle cavity, may fold over the rim of the shell
and extend over part of it. In parasitic gastropods the shell may be absent and the adult
somewhat worm-like, however, the juvenile forms still possess a typical spiral shell. If you hold
a shell with the aperture facing you and the apex pointing up, then you will discover that most
shells are right-handed or dextral, that is the aperture is on the right. However, left-handed
or sinistral shells are normal for some species.
The shell consists of an outer organic layer of sclerotised (hardened) protein called
chonchin or chonchiolin. This thin layer is called the periostracum. This layer may erode
and be absent in many mature shells and is lacking in those where the mantle covers the
outer surface of the shell, as in cowries. Beneath this are (usually) three layers comprised
mostly of calcium carbonate: the outermost is the prismatic or pallisade layer (consists of
layers perpendicular to the shell surface), the middle is the lamellated layer (consists of layers
parallel to the shell surface) and the innermost is the nacreous layer (nacreous means
pearly). Terrestrial pulmonates (snails that live on land and do not have gills) have 4 to 6
calcareous layers in their shells.
Of the different crystalline forms of calcium carbonate, calcite occurs in the reduced shells of
naked land slugs and pelagic sea-snails (heteropods). Aragonite occurs in many
prosobranchs, most pulmonates (land snails) and some sea slugs (opsithobranchs).
Aragonite is less stable and reverts to calcite in fossils. Some shells are a mixture of the two
forms. Other calcium slats may be present, such as calcium phosphate and calcium sulphate
and some magneisum salts. Sometimes the calcareous layers are interlaced with conchin.
The Gastropod Body
The body of the gastropod is divided into head, foot, mantle and visceral mass. There is a
definite head and the animals have bilateral symmetry. There are cephalic tentacles (head
tentacles) of some sort, eyes and a terminal mouth. The ventral body forms a muscular foot
which forms a flat creeping sole. It may carry a shield on its back called the operculum,
which plugs the aperture of the shell when the snail is withdrawn into its shell. The mantle is a
collarlike fold of the body wall which lines the body whorl and encloses the mantle cavity
(pallial cavity) between itself and the protrusible part of the body (head and foot). The mantle
may be drawn out into a tube called the siphon, which lies in the siphonal canal of the shell.
On the mantle cavity is the pallial complex, consisting of a single gill (ctenidium) or a pair of
gills, anus, nephridiopore, gonopore (usually single, but may be paired), one or two
hypobranchial gland, one or two osphradia (sing. osphradium) and the terminal parts of the
digestive, excretory and reproductive systems behind their respective openings. In most the
right pair of these organs (gills, osphradia etc.) is generally missing (originally this would have
been the left side pre-torsion).
The hypobranchial gland is a glandular region of epidermis, which may be thrown into a
series of folds at right-angles to the mantle wall, lining the mantle roof (sometimes there is no
definite glandular area). When both are present, then the left is larger than the right. This
contains mucocytes, which secrete lubricating mucus in response to irritants, such as sand,
caught in the mantle cavity. The dye known as Tyrian purple is made from the hypobranchial
secretions of certain Murex species.
The skin of the gastropod consists of a single layer of epithelial cells, usually covered
externally by a secreted cuticle. This epithelium is often ciliated.The osphradium resembles a
miniature gill adjacent to the real gill, and may function as a chemoreceptive organ (organ of
smell / taste).
Interiorly, there is no coelom around the viscera (as there is in vertebrates, for example).
There are blood spaces (sinuses), fibrous connective tissue and muscle fibres. The interior
may be gelatinous. Star-shaped (stellate) cells with anastomosing processes, amoeboid cells
and clear chondroid cells may be scattered throughout the connective tissue. Calcareous
inclusions may be present inside cells or outside cells embedded in the tissues. Pigment cells
may also be present.
The visceral mass is permanently contained in the larger turns of the spire and is often coiled
and contains most of the digestive tract, the heart, the nephridium (excretory organ or
'kidney') and much of the reproductive system. In terrestrial slugs and some shell-less sea
slugs (nudibranchs) the visceral mass and foot are not distinctly marked off from one-another
but form a single mass.
head with a pair of eyes, cephalic tentacles and a proboscis. To the animal's right is the large
siphon, whilst to the left is the penis tucked away inside the shell. Notice the operculum or
'shield' on the back of the foot.
Sea Snails
The gastropods are molluscs and so are included in the same phylum as cephalopods and
bivalves. The name 'gastropod' literally means 'stomach-foot' in reference to the way snails
and slugs appear to crawl on their bellies. In reality the stomach and other parts of the gut are
contained in the dorsal hump (beneath the protective shell in snails). Most snails belong to a
group of the gastropods referred to as prosobranchs. Prosobranchs are a diverse group
with multiple ancestors, and so gastropod classification has been overhauled in recent years
to reflect evolutionary relationships. Though useful to studies of evolution, evolutionary
classifications are of limited field use and we shall stick only to descriptive terms in this article,
avoiding complex issues of 'family trees'.
The prosobranchs are 'fore-gilled' snails. What does this mean? Well, in ancestral forms the
anus and pair of excretory pores (nephridiopores) opened at the rear in the mantle cavity.
The mantle cavity is an external space just underneath the rim of the shell, lined of course
by the epithelium (or 'skin') of the snail. A pair of external gills (borne of the surface of the
snail's body) also opened into the rear of this mantle cavity. Thus the gills are in a protected
cavity beneath the shell but open to sea water in the mantle cavity. During the course of
evolution, the gastropod snails underwent torsion. In the developing snail the hump, with its
shell and internal organs, twists through 180 degrees, so that the anus, excretory pores and
gills now sit above the head in the anterior mantle cavity.
The Gastropod Shell
There are some 85 000 described species of snail, though this probably represents a small
fraction. Most are marine, some are freshwater and some are terrestrial. The shell consists of
a cone wound into a spiral about a central axis or columella. Each turn of the spiral is a
whorl and whorls are distinguishable in most cases by external grooves or lines in the shell
called sutures, which may be simple curves or wavy boundaries. The basal whorl is the
largest and called the body whorl and its boundary is the open aperture through which the
foot and head can be protruded or retracted. This aperture defines the anterior or head-end
of the shell, whilst the apex of the shell is posterior. Delimiting the aperture is an outer lip of
shell opposite the inner lip, the latter is continuous with the columella. The smaller (and
older) whorls of the shell form the spire, which decreases in size towards the pointed apex.
The shell surface may be smooth or sculptured with ridges, grooves, ribs, tubercles or spines.
Many marine forms are brightly coloured with spiral bands, blotches, stripes or dots. These
materials may be deposited waste products, or they may have a camouflage function and
include melanins, pyrroles and porphyrins.
The shell may bear a siphonal canal at the front through which the siphon (if present) can
irrigate the mantle cavity with sea water to facilitate gas exchange at the gills and the ejection
of waste. The siphonal canal is an extension of the body whorl and aperture. Opposite there
may be a similar and smaller posterior canal.
The shell may be long and slender with many whorls or short and plump with a reduced spire.
The spire may also be incorporated into the body whorl and so not readily apparent. All the
whorls may lie in one plane (a planospiral shell) or it may be helical. Limpets have conical
shells with no spire and an aperture that occupies the whole width of the shell. In cowries, the
aperture is reduced to an elongated slit.
The mantle, which is the soft tissue lining the mantle cavity, may fold over the rim of the shell
and extend over part of it. In parasitic gastropods the shell may be absent and the adult
somewhat worm-like, however, the juvenile forms still possess a typical spiral shell. If you hold
a shell with the aperture facing you and the apex pointing up, then you will discover that most
shells are right-handed or dextral, that is the aperture is on the right. However, left-handed
or sinistral shells are normal for some species.
The shell consists of an outer organic layer of sclerotised (hardened) protein called
chonchin or chonchiolin. This thin layer is called the periostracum. This layer may erode
and be absent in many mature shells and is lacking in those where the mantle covers the
outer surface of the shell, as in cowries. Beneath this are (usually) three layers comprised
mostly of calcium carbonate: the outermost is the prismatic or pallisade layer (consists of
layers perpendicular to the shell surface), the middle is the lamellated layer (consists of layers
parallel to the shell surface) and the innermost is the nacreous layer (nacreous means
pearly). Terrestrial pulmonates (snails that live on land and do not have gills) have 4 to 6
calcareous layers in their shells.
Of the different crystalline forms of calcium carbonate, calcite occurs in the reduced shells of
naked land slugs and pelagic sea-snails (heteropods). Aragonite occurs in many
prosobranchs, most pulmonates (land snails) and some sea slugs (opsithobranchs).
Aragonite is less stable and reverts to calcite in fossils. Some shells are a mixture of the two
forms. Other calcium slats may be present, such as calcium phosphate and calcium sulphate
and some magneisum salts. Sometimes the calcareous layers are interlaced with conchin.
The Gastropod Body
The body of the gastropod is divided into head, foot, mantle and visceral mass. There is a
definite head and the animals have bilateral symmetry. There are cephalic tentacles (head
tentacles) of some sort, eyes and a terminal mouth. The ventral body forms a muscular foot
which forms a flat creeping sole. It may carry a shield on its back called the operculum,
which plugs the aperture of the shell when the snail is withdrawn into its shell. The mantle is a
collarlike fold of the body wall which lines the body whorl and encloses the mantle cavity
(pallial cavity) between itself and the protrusible part of the body (head and foot). The mantle
may be drawn out into a tube called the siphon, which lies in the siphonal canal of the shell.
On the mantle cavity is the pallial complex, consisting of a single gill (ctenidium) or a pair of
gills, anus, nephridiopore, gonopore (usually single, but may be paired), one or two
hypobranchial gland, one or two osphradia (sing. osphradium) and the terminal parts of the
digestive, excretory and reproductive systems behind their respective openings. In most the
right pair of these organs (gills, osphradia etc.) is generally missing (originally this would have
been the left side pre-torsion).
The hypobranchial gland is a glandular region of epidermis, which may be thrown into a
series of folds at right-angles to the mantle wall, lining the mantle roof (sometimes there is no
definite glandular area). When both are present, then the left is larger than the right. This
contains mucocytes, which secrete lubricating mucus in response to irritants, such as sand,
caught in the mantle cavity. The dye known as Tyrian purple is made from the hypobranchial
secretions of certain Murex species.
The skin of the gastropod consists of a single layer of epithelial cells, usually covered
externally by a secreted cuticle. This epithelium is often ciliated.The osphradium resembles a
miniature gill adjacent to the real gill, and may function as a chemoreceptive organ (organ of
smell / taste).
Interiorly, there is no coelom around the viscera (as there is in vertebrates, for example).
There are blood spaces (sinuses), fibrous connective tissue and muscle fibres. The interior
may be gelatinous. Star-shaped (stellate) cells with anastomosing processes, amoeboid cells
and clear chondroid cells may be scattered throughout the connective tissue. Calcareous
inclusions may be present inside cells or outside cells embedded in the tissues. Pigment cells
may also be present.
The visceral mass is permanently contained in the larger turns of the spire and is often coiled
and contains most of the digestive tract, the heart, the nephridium (excretory organ or
'kidney') and much of the reproductive system. In terrestrial slugs and some shell-less sea
slugs (nudibranchs) the visceral mass and foot are not distinctly marked off from one-another
but form a single mass.
Sea
Slugs
Sea slugs are an incredibly diverse and beautiful group of gastropods! Many sea slugs are
also known as nudibranchs, literally 'naked gills'. However, some creatures less commonly
called 'sea slugs' belong to a few other groups of gastropods, but all these and the
nudibranchs belong to a group called the opisthobranchs. This includes the sea hares
(aplysiomorphs), sea butterflies (pteropods) and sea angels (Gymnosomata). In nudibranchs,
the mantle has overgrown the shell which has disappeared. In some land slugs, the mantle
has similarly overgrown the shell, but the shell persists as an internal operculum.
Sea slugs are an incredibly diverse and beautiful group of gastropods! Many sea slugs are
also known as nudibranchs, literally 'naked gills'. However, some creatures less commonly
called 'sea slugs' belong to a few other groups of gastropods, but all these and the
nudibranchs belong to a group called the opisthobranchs. This includes the sea hares
(aplysiomorphs), sea butterflies (pteropods) and sea angels (Gymnosomata). In nudibranchs,
the mantle has overgrown the shell which has disappeared. In some land slugs, the mantle
has similarly overgrown the shell, but the shell persists as an internal operculum.
Respiration
The gill or ctenidium consists of a longitudinal series of thin leaves borne on a central axis.
Ancestrally there was a symmetrically placed pair of gills, each with a double row of leaves
(bipectinate), but in present forms there is often one gill with one row of leaves
(monopectinate). The gill is supplied by afferent (incoming) and efferent (outgoing) blood
sinuses. Each leaf or leaflet is triangular, supported by skeletal rods, and with cilia along its
margins. Incoming water flows over one side of the leaflet, outgoing water over the other.
Incoming water enters the siphon or spout, if present (which is usually on the left side) and
passes over the gills in the mantle cavity and exits, usually at the right side of the snail's head,
or via holes in the shell as in keyhole limpets and Haliotis (abalone). Heavy particles caught in
the stream are rejected near the entrance.
Many land snails (pulmonates) have no gills, instead the mantle edge fuses with the neck and
a small hole opens into this mantle cavity which is now the pulmonary sac. In this case the
mantle roof is highly vascularised for gas exchange. Nudibranchs have no mantle cavity and in
place of the standard gills they have finger-like, feather-like or variously lobed projections
called cerata. Some gastropods have no gills or other specialised respiratory organs. Some
amphibious snails have both a gill and a pulmonary sac and cans witch from one to the other.
Some snails can survive being buried in mud for months (e.g. during a dry season) by storing
oxygen in their pulmonary sac or by respiring anaerobically.
Circulation
The heart is enclosed in a pericardial cavity and has only one ventricle. In forms with two gills
there are two atria, but in forms with a single gill there is usually only one atrium. The heart is
a more-or-less straight tube. It does not require the elaborate coiling of the vertebrate heart
since its blood is flowing under low pressure and it does not need to capitalise on the
momentum of the flowing blood to conserve energy. (In the mammal, for example, the heart is
twisted to follow the tendency for blood to spin round when entering the chambers at high
speed, so that the heart pumps with the blood rather than against it). Vessels enter and leave
the gastropod heart, but the system is an open circulation since blood exits the outgoing
vessels to flow through sinuses (blood spaces). Blood is delivered to the gills (and back to the
heart) and to the nephridium.
Excretion
Ancestrally a pair of metanephridia occurred close to the pericardium, but in forms with one gill
and one atrium, there is only one nephridium. The metanephridium is permeated with blood
returning to the heart and opens in the pericardial cavity and via a nephridiopore in the mantle
cavity through which metabolic wastes are ejected.
Nervous System
Definite ganglia or nerve centres occur in groups around the pharynx. There is one pair of
cerebral ganglia above the pharynx, one pair of pleural or pallial ganglia at the sides of the
pharynx and one pair of pedal gangia below the pharynx. There is also a visceral ganglion
connected to the pleural ganglia by pleurovisceral connectives, forming a figure of eight (with
one connective below and one above the foregut) - an arrangement resulting from torsion.
The cerebral, pleural and pedal ganglia together with the commissures (tracts) connecting
them form a circumenteric ring (essentially a ring-like 'brain' around the pharynx). The
cerebral ganglia give off a tentacular nerve to each tentacle and an optic nerve to each
eye and a statocyst nerve to each statocyst (see 'sensory organs' below). Either the
cerebral ganglia or the cerebral commissure or nerve tract which connects them give out
labial nerves to the lips, snout and anterior end and dorsal wall of the head. The pleural
ganglia (which may be fused to the pedal ganglia) supply the mantle and columellar muscle
with nerves. The intestinal ganglia give out one pair of visceral nerves to connect to the
visceral (abdominal) ganglion at the beginning of the visceral mass. The intestinal and visceral
ganglia innervate the gills (sometimes via branchial ganglia) and osphradium. From the
visceral ganglion, the rectal nerve innervates the visceral mass (heart, nephridium,
pericardium, reproductive system and postoesophageal digetsive tract).
Sensory Organs
Neurosensory cells occur in the epdiermis covering the gastropod's body and these may be
grouped into sensory buds. These are also found on the vraious papillae and tentacles which
may be present, such as cephalic or pedal tentacles, or papillae on the mantle. Some have
one or a pair of osphradia (singular: osphradium) situated on the underside of the mantle
roof 9in the pallial complex) or base of the siphon. The osphradium resembles a miniature
bipectinate gill or there may be a row of warts or row of ciliate cells or thickened epithelium
near each gill. Neurosensory cells may be found here and this organ is probably
chemosensory (olfactory), sensing and testing the inflowing water.
A pair of statocysts are often found in the foot. These are innervated by the cerebral ganglia
and each consists of a spherical or oval fluid-filled vesicle with one large statolith (stony
structure) or several small statoconia anchored to the statocyst wall. These presumably sense
changes in gravity relative to the gastropod. The highly-mobile and pelagic heteropods have
large and especially complicated statoliths, whilst sessile forms that move little have none.
A pair of eyes is usually present; each eye borne on the outer side of a cephalic tentacle,
either on the tentacle itself, at the tentacle base or borne on its own stalk. The eyes may be
absent or reduced in cave-dwelling forms, abyssal forms and some burrowing forms. The eye
may consist of a simple pit lined by photoreceptors, but in some forms the pit closes over to
form a vesicle complete with a lens. The pelagic heteropods have especially complex eyes,
with a lens, cornea and retina and muscles to move them.
Reproduction
Prosobranchs are mostly dioecious; opisthobranchs and pulmonates are hermaphroditic, with
one hermaphroditic gonad and a gonoduct, with various glandular appendages, that opens
via the gonopore or nephridiopore. The sperm duct of the male may open as a pore or ciliated
groove in the mantle cavity or via a penis, which is usually on the right side of the head or
neck. The penis is muscular but also has a ciliated lumen, suggesting that cilia action is
important for transport of sperm to the opening. Alternatively the right tentacle may act as an
intromittent organ. Many produce two types of spermatozoa. The fertile flagellate (eupyrene
sperm and sterile vermiform (apyrene0 spermatozoa. The normal sperm may surround and
adhere to the vermiform type, which may be supplying the spermatozoa with nutrients. Some
release the sperm in packets called spermatophores which is inserted in the female (after the
sperm are utilised, the female digests the empty case). In forms with external fertilisation, the
excretory nephridium may discharge the sex cells through the nephridiopore on the right side
of the animal. Heteropod males use a penis to deposit spermatophores on the female.
The female systems are very variable, and only a basic outline of the main types will be given
here. The female system opens via the oviduct in the mantle roof, near the anus, via the
female gonopore or via the nephrdium and nephridiopore on the right side. A protrusible
ovipositor may be present and even a rudimentary penis in the female. In those forms with
internal fertilisation, seminal receptacles receive the sperm from the male, either via the
gonopore or a pore adjacent to it. The sperm enter the bursa copulatrix, which is a blind
pouch (a duct or vagina may connect the exterior pore to the bursa). The sperm are then
transferred to the seminal receptacle(s) by direct connection or via a ciliated channel in the
oviduct. The bursa may digest excess sperm. A crystal sac may connect to the main
gonopore. This contains foreign particles, such as sand grains, which can be added to the
developing egg shell.
The slipper limpet, Crepidula, is a protandric hermaphrodite. This means that the individual is
initially male. These limpets stack on top of one-another for reproduction and the individual at
the bottom of the stack becomes a female. Up to 18 males may be stacked on top of this
single female. The female produces a pheromone which attracts young limpets to settle on top
of her, at which point they develop into males. If a male is removed then it differentiates
irreversibly into a female. Crepidula may spawn up to ten times a year, producing 50 to 100
egg capsules, each containing 100 to 200 eggs! The egg capsules are brooded beneath the
foot and spawning does not occur whilst egg capsules are being brooded.
Some species are hermaphrodites and self-fertilisation is possible in some. In others males
are rare or possibly completely absent, in which case reproduction is presumably by
parthenogenesis.
Coastal forms may synchronise spawning in accordance with the lunar cycle, e.g. Littorina
littorea. Some shed their sex cells into the sea, where they are fertilized. Haliotis sheds sperm
and eggs into the sea via its respiratory pore. In others, fertilisation is internal. Egg capsules
of some species may be found strewn across mud flats, others are attached to the parent, or
deposited on other organisms or the eggs may float. The egg capsules may have protein
plugs which dissolve at hatching, or cemented lids in which the cement dissolves. Most eggs
do not hatch and possibly act as food for the hatchlings. Eggs may be brooded in a uterus
(modified part of the oviduct), in the mantle cavity or in a pouch on the body surface.
The fertilised egg undergoes spiral cleavage which is also detrminate. After gastrulation, the
fertilized egg develops into a trochophore larva. (See polychaetes for more on trochophore
larvae). These larvae are free-swimming and planktonic. Most, however, copulate and
fertilization is internal and eggs are laid in gelatinous strings or masses or enclosed in
secreted capsules. Strombus lays gelatinous strings containing up to 460 000 eggs and these
strings may be 50 to 75 feet (about 25 m) in length! In fresh-water and terrestrial snails, baby
snails hatch, and also in viviparous or ovoviviparous marine snails. In other marine snails, the
fertilized egg develops into a veliger larva. The velliger has a bilobed (sometimes 3, 4 or
6-lobed) ciliated velum which is carried in front and a shell into which the velum can be
withdrawn. The velliger has a larval heart (which does not develop into the adult heart) and an
open circulation. Blood circulates through the velar lobes. There is no excretory
protonephridium. The velliger is a ciliary feeder, trapping diatoms and other small food items
in currents generated by the beating cilia on the velum. A food groove on the velum carries
the trapped particles of food to the mouth.
The velliger larva undergoes torsion (the shell and visceral mass rotate 180 degrees
counterclockwise) after several minutes or hours and lasts for hours or 2-4 weeks and is
planktonic. It eventually metamorphoses into a young snail. The velum is shed, the larval heart
regresses and the velliger sinks to the bottom. The larval shell is either retained or cast off.
The adults may live up to about 20 years in some species.
Regeneration
The shell can repair holes and lost tentacles and eyes can be regenerated in some species
over a period of weeks or months. The entire foot and the siphon can also be regenerated in
some forms. The blood clots and wounds close over.
Locomotion
Small species may glide by means of beating cilia on the bottom of the foot or sole. Larger
forms utilise muscular waves which travel along the foot. During forwards locomotion, the
waves may pass from the posterior to the anterior of the foot or from anterior to posterior
(retrograde). Speeds of up to 70 cm / minute may be attained. All forms, except limpets, seem
able to right themselves. Some forms float, by secreting mucus full of air bubbles to form a
float attached to the rear part of the foot (metapodium) as in the Violet Snails (Lanthina, a
mesogastropod). Some swim by thrashing the front part of the foot (propodium) as in
Polinices. Heteropods swim by means of the flattened propodium. In Bullia and Terebra, the
snails float on their side with the foot expanded as a snail.
Classification
The classification of gastropods has undergone extensive revision in an attempt to make it
better reflect genetic and evolutionary relationships. Most field guides, however, still use the
old classification and the groups used in this division are still used as informal groupings and
descriptive terms for groups which may have several evolutionary origins. Prosobranchs
contain the archaeogastropods, the mesogastropods and the neogastropods.
The archaeogastropods often still have two bipectinate gills and two nephridia. The gonad
discharges through the right nephridium and nephridiopore. They lack a siphon, proboscis
and penis. Examples include the Ormer (Haliotis), keyhole limpets (Diodora) and totoiseshell
limpets (Acmaea), limpets (Patella), topshells (Gibbula and Monodonta) and many others.
The mesogastropods have one monopectinate ctenidium (gill) and one nephridium. The
shell is usually conical. Examples include the periwinkles (Littorina), tower shells (Turritella),
the Pelican's foot shell (Aporrhais), the slipper limpet (Crepidula) and cowries.
The neogastropods have well-developed siphons which are often supported by a siphonal
groove or canal in the shell. Many are carnivorous. Examples include the common whelk
(Buccinum), the dogwhelk (Nucella), the Oyster drill (Ocenebra), Murex and cone shells
(Conus).
Ecology
Gastropods are found in all seas and at all latitudes, from the intertidal zone down to 8200 m.
They are primarily benthonic, but pteropods and heteropods secondarily evolved as swimmers
and the janthinids secondarily evolved a floating lifestyle. Many forms are fresh-water and
terrestrial. Terrestrial forms are not found on glaciers and alpine summits, but are found
elsewhere. Some forms are parasitic on echinoderms and have snail-like young. Some are
attached and sessile. There are three subclasses: Prosobranchia, Opisthobranchia and
Pulmonata.
The gill or ctenidium consists of a longitudinal series of thin leaves borne on a central axis.
Ancestrally there was a symmetrically placed pair of gills, each with a double row of leaves
(bipectinate), but in present forms there is often one gill with one row of leaves
(monopectinate). The gill is supplied by afferent (incoming) and efferent (outgoing) blood
sinuses. Each leaf or leaflet is triangular, supported by skeletal rods, and with cilia along its
margins. Incoming water flows over one side of the leaflet, outgoing water over the other.
Incoming water enters the siphon or spout, if present (which is usually on the left side) and
passes over the gills in the mantle cavity and exits, usually at the right side of the snail's head,
or via holes in the shell as in keyhole limpets and Haliotis (abalone). Heavy particles caught in
the stream are rejected near the entrance.
Many land snails (pulmonates) have no gills, instead the mantle edge fuses with the neck and
a small hole opens into this mantle cavity which is now the pulmonary sac. In this case the
mantle roof is highly vascularised for gas exchange. Nudibranchs have no mantle cavity and in
place of the standard gills they have finger-like, feather-like or variously lobed projections
called cerata. Some gastropods have no gills or other specialised respiratory organs. Some
amphibious snails have both a gill and a pulmonary sac and cans witch from one to the other.
Some snails can survive being buried in mud for months (e.g. during a dry season) by storing
oxygen in their pulmonary sac or by respiring anaerobically.
Circulation
The heart is enclosed in a pericardial cavity and has only one ventricle. In forms with two gills
there are two atria, but in forms with a single gill there is usually only one atrium. The heart is
a more-or-less straight tube. It does not require the elaborate coiling of the vertebrate heart
since its blood is flowing under low pressure and it does not need to capitalise on the
momentum of the flowing blood to conserve energy. (In the mammal, for example, the heart is
twisted to follow the tendency for blood to spin round when entering the chambers at high
speed, so that the heart pumps with the blood rather than against it). Vessels enter and leave
the gastropod heart, but the system is an open circulation since blood exits the outgoing
vessels to flow through sinuses (blood spaces). Blood is delivered to the gills (and back to the
heart) and to the nephridium.
Excretion
Ancestrally a pair of metanephridia occurred close to the pericardium, but in forms with one gill
and one atrium, there is only one nephridium. The metanephridium is permeated with blood
returning to the heart and opens in the pericardial cavity and via a nephridiopore in the mantle
cavity through which metabolic wastes are ejected.
Nervous System
Definite ganglia or nerve centres occur in groups around the pharynx. There is one pair of
cerebral ganglia above the pharynx, one pair of pleural or pallial ganglia at the sides of the
pharynx and one pair of pedal gangia below the pharynx. There is also a visceral ganglion
connected to the pleural ganglia by pleurovisceral connectives, forming a figure of eight (with
one connective below and one above the foregut) - an arrangement resulting from torsion.
The cerebral, pleural and pedal ganglia together with the commissures (tracts) connecting
them form a circumenteric ring (essentially a ring-like 'brain' around the pharynx). The
cerebral ganglia give off a tentacular nerve to each tentacle and an optic nerve to each
eye and a statocyst nerve to each statocyst (see 'sensory organs' below). Either the
cerebral ganglia or the cerebral commissure or nerve tract which connects them give out
labial nerves to the lips, snout and anterior end and dorsal wall of the head. The pleural
ganglia (which may be fused to the pedal ganglia) supply the mantle and columellar muscle
with nerves. The intestinal ganglia give out one pair of visceral nerves to connect to the
visceral (abdominal) ganglion at the beginning of the visceral mass. The intestinal and visceral
ganglia innervate the gills (sometimes via branchial ganglia) and osphradium. From the
visceral ganglion, the rectal nerve innervates the visceral mass (heart, nephridium,
pericardium, reproductive system and postoesophageal digetsive tract).
Sensory Organs
Neurosensory cells occur in the epdiermis covering the gastropod's body and these may be
grouped into sensory buds. These are also found on the vraious papillae and tentacles which
may be present, such as cephalic or pedal tentacles, or papillae on the mantle. Some have
one or a pair of osphradia (singular: osphradium) situated on the underside of the mantle
roof 9in the pallial complex) or base of the siphon. The osphradium resembles a miniature
bipectinate gill or there may be a row of warts or row of ciliate cells or thickened epithelium
near each gill. Neurosensory cells may be found here and this organ is probably
chemosensory (olfactory), sensing and testing the inflowing water.
A pair of statocysts are often found in the foot. These are innervated by the cerebral ganglia
and each consists of a spherical or oval fluid-filled vesicle with one large statolith (stony
structure) or several small statoconia anchored to the statocyst wall. These presumably sense
changes in gravity relative to the gastropod. The highly-mobile and pelagic heteropods have
large and especially complicated statoliths, whilst sessile forms that move little have none.
A pair of eyes is usually present; each eye borne on the outer side of a cephalic tentacle,
either on the tentacle itself, at the tentacle base or borne on its own stalk. The eyes may be
absent or reduced in cave-dwelling forms, abyssal forms and some burrowing forms. The eye
may consist of a simple pit lined by photoreceptors, but in some forms the pit closes over to
form a vesicle complete with a lens. The pelagic heteropods have especially complex eyes,
with a lens, cornea and retina and muscles to move them.
Reproduction
Prosobranchs are mostly dioecious; opisthobranchs and pulmonates are hermaphroditic, with
one hermaphroditic gonad and a gonoduct, with various glandular appendages, that opens
via the gonopore or nephridiopore. The sperm duct of the male may open as a pore or ciliated
groove in the mantle cavity or via a penis, which is usually on the right side of the head or
neck. The penis is muscular but also has a ciliated lumen, suggesting that cilia action is
important for transport of sperm to the opening. Alternatively the right tentacle may act as an
intromittent organ. Many produce two types of spermatozoa. The fertile flagellate (eupyrene
sperm and sterile vermiform (apyrene0 spermatozoa. The normal sperm may surround and
adhere to the vermiform type, which may be supplying the spermatozoa with nutrients. Some
release the sperm in packets called spermatophores which is inserted in the female (after the
sperm are utilised, the female digests the empty case). In forms with external fertilisation, the
excretory nephridium may discharge the sex cells through the nephridiopore on the right side
of the animal. Heteropod males use a penis to deposit spermatophores on the female.
The female systems are very variable, and only a basic outline of the main types will be given
here. The female system opens via the oviduct in the mantle roof, near the anus, via the
female gonopore or via the nephrdium and nephridiopore on the right side. A protrusible
ovipositor may be present and even a rudimentary penis in the female. In those forms with
internal fertilisation, seminal receptacles receive the sperm from the male, either via the
gonopore or a pore adjacent to it. The sperm enter the bursa copulatrix, which is a blind
pouch (a duct or vagina may connect the exterior pore to the bursa). The sperm are then
transferred to the seminal receptacle(s) by direct connection or via a ciliated channel in the
oviduct. The bursa may digest excess sperm. A crystal sac may connect to the main
gonopore. This contains foreign particles, such as sand grains, which can be added to the
developing egg shell.
The slipper limpet, Crepidula, is a protandric hermaphrodite. This means that the individual is
initially male. These limpets stack on top of one-another for reproduction and the individual at
the bottom of the stack becomes a female. Up to 18 males may be stacked on top of this
single female. The female produces a pheromone which attracts young limpets to settle on top
of her, at which point they develop into males. If a male is removed then it differentiates
irreversibly into a female. Crepidula may spawn up to ten times a year, producing 50 to 100
egg capsules, each containing 100 to 200 eggs! The egg capsules are brooded beneath the
foot and spawning does not occur whilst egg capsules are being brooded.
Some species are hermaphrodites and self-fertilisation is possible in some. In others males
are rare or possibly completely absent, in which case reproduction is presumably by
parthenogenesis.
Coastal forms may synchronise spawning in accordance with the lunar cycle, e.g. Littorina
littorea. Some shed their sex cells into the sea, where they are fertilized. Haliotis sheds sperm
and eggs into the sea via its respiratory pore. In others, fertilisation is internal. Egg capsules
of some species may be found strewn across mud flats, others are attached to the parent, or
deposited on other organisms or the eggs may float. The egg capsules may have protein
plugs which dissolve at hatching, or cemented lids in which the cement dissolves. Most eggs
do not hatch and possibly act as food for the hatchlings. Eggs may be brooded in a uterus
(modified part of the oviduct), in the mantle cavity or in a pouch on the body surface.
The fertilised egg undergoes spiral cleavage which is also detrminate. After gastrulation, the
fertilized egg develops into a trochophore larva. (See polychaetes for more on trochophore
larvae). These larvae are free-swimming and planktonic. Most, however, copulate and
fertilization is internal and eggs are laid in gelatinous strings or masses or enclosed in
secreted capsules. Strombus lays gelatinous strings containing up to 460 000 eggs and these
strings may be 50 to 75 feet (about 25 m) in length! In fresh-water and terrestrial snails, baby
snails hatch, and also in viviparous or ovoviviparous marine snails. In other marine snails, the
fertilized egg develops into a veliger larva. The velliger has a bilobed (sometimes 3, 4 or
6-lobed) ciliated velum which is carried in front and a shell into which the velum can be
withdrawn. The velliger has a larval heart (which does not develop into the adult heart) and an
open circulation. Blood circulates through the velar lobes. There is no excretory
protonephridium. The velliger is a ciliary feeder, trapping diatoms and other small food items
in currents generated by the beating cilia on the velum. A food groove on the velum carries
the trapped particles of food to the mouth.
The velliger larva undergoes torsion (the shell and visceral mass rotate 180 degrees
counterclockwise) after several minutes or hours and lasts for hours or 2-4 weeks and is
planktonic. It eventually metamorphoses into a young snail. The velum is shed, the larval heart
regresses and the velliger sinks to the bottom. The larval shell is either retained or cast off.
The adults may live up to about 20 years in some species.
Regeneration
The shell can repair holes and lost tentacles and eyes can be regenerated in some species
over a period of weeks or months. The entire foot and the siphon can also be regenerated in
some forms. The blood clots and wounds close over.
Locomotion
Small species may glide by means of beating cilia on the bottom of the foot or sole. Larger
forms utilise muscular waves which travel along the foot. During forwards locomotion, the
waves may pass from the posterior to the anterior of the foot or from anterior to posterior
(retrograde). Speeds of up to 70 cm / minute may be attained. All forms, except limpets, seem
able to right themselves. Some forms float, by secreting mucus full of air bubbles to form a
float attached to the rear part of the foot (metapodium) as in the Violet Snails (Lanthina, a
mesogastropod). Some swim by thrashing the front part of the foot (propodium) as in
Polinices. Heteropods swim by means of the flattened propodium. In Bullia and Terebra, the
snails float on their side with the foot expanded as a snail.
Classification
The classification of gastropods has undergone extensive revision in an attempt to make it
better reflect genetic and evolutionary relationships. Most field guides, however, still use the
old classification and the groups used in this division are still used as informal groupings and
descriptive terms for groups which may have several evolutionary origins. Prosobranchs
contain the archaeogastropods, the mesogastropods and the neogastropods.
The archaeogastropods often still have two bipectinate gills and two nephridia. The gonad
discharges through the right nephridium and nephridiopore. They lack a siphon, proboscis
and penis. Examples include the Ormer (Haliotis), keyhole limpets (Diodora) and totoiseshell
limpets (Acmaea), limpets (Patella), topshells (Gibbula and Monodonta) and many others.
The mesogastropods have one monopectinate ctenidium (gill) and one nephridium. The
shell is usually conical. Examples include the periwinkles (Littorina), tower shells (Turritella),
the Pelican's foot shell (Aporrhais), the slipper limpet (Crepidula) and cowries.
The neogastropods have well-developed siphons which are often supported by a siphonal
groove or canal in the shell. Many are carnivorous. Examples include the common whelk
(Buccinum), the dogwhelk (Nucella), the Oyster drill (Ocenebra), Murex and cone shells
(Conus).
Ecology
Gastropods are found in all seas and at all latitudes, from the intertidal zone down to 8200 m.
They are primarily benthonic, but pteropods and heteropods secondarily evolved as swimmers
and the janthinids secondarily evolved a floating lifestyle. Many forms are fresh-water and
terrestrial. Terrestrial forms are not found on glaciers and alpine summits, but are found
elsewhere. Some forms are parasitic on echinoderms and have snail-like young. Some are
attached and sessile. There are three subclasses: Prosobranchia, Opisthobranchia and
Pulmonata.


Left:
the radula of Pila. The gastropod
radula consists of a supporting subradula
membrane, consisting of chitin, epithelial
cells and connective tissue, and rows of
teeth. There is a column of median
teeth (middle, axial or rachidian teeth)
with one or more lateral teeth on either
side, the number depending on species.
There may also be marginal teeth or
uncini right on the edge of the radula.
There may be as many as four different
tooth types on the same radula, including
bristle-bearing teeth. In the example
shown here, there are only three teeth
per row, but the median teeth have
several cusps (points). Thus we have a
(1.1.1) arrangement - one medial tooth
with one lateral tooth either side of it.
Pila is a freshwater snail belonging to a
group of gastropods called the
neogastropods, which includes the
whelks (Buccinum), dogwhelks, Murex,
Conus (cone shells). The members of
this group have three teeth per row of
the radula and many are predators. The
radula of cone shells is modified into a
protrusible venemous harpoon on the tip
of its proboscis. This radula has two rows
of teeth with pointed (lanceolate) tips.
Another gastropod group, the
mesogatropods, includes the periwinkles
(Littorina), tower shells and worm shells.
These gastropods have seven teeth per
row of the radula, giving the formula:
(2.1.1.1.2) for one pair of laterals and
two pairs of marginals. The heteropods,
a group of pelagic (free
swimming/floating) snails, are also
included in this group and in these the
marginal teeth are spinelike and used for
grasping prey.
The radula is generally absent from
parasitic forms.
radula consists of a supporting subradula
membrane, consisting of chitin, epithelial
cells and connective tissue, and rows of
teeth. There is a column of median
teeth (middle, axial or rachidian teeth)
with one or more lateral teeth on either
side, the number depending on species.
There may also be marginal teeth or
uncini right on the edge of the radula.
There may be as many as four different
tooth types on the same radula, including
bristle-bearing teeth. In the example
shown here, there are only three teeth
per row, but the median teeth have
several cusps (points). Thus we have a
(1.1.1) arrangement - one medial tooth
with one lateral tooth either side of it.
Pila is a freshwater snail belonging to a
group of gastropods called the
neogastropods, which includes the
whelks (Buccinum), dogwhelks, Murex,
Conus (cone shells). The members of
this group have three teeth per row of
the radula and many are predators. The
radula of cone shells is modified into a
protrusible venemous harpoon on the tip
of its proboscis. This radula has two rows
of teeth with pointed (lanceolate) tips.
Another gastropod group, the
mesogatropods, includes the periwinkles
(Littorina), tower shells and worm shells.
These gastropods have seven teeth per
row of the radula, giving the formula:
(2.1.1.1.2) for one pair of laterals and
two pairs of marginals. The heteropods,
a group of pelagic (free
swimming/floating) snails, are also
included in this group and in these the
marginal teeth are spinelike and used for
grasping prey.
The radula is generally absent from
parasitic forms.
Digestive
Tract and Nutrition
The mouth, which is often borne on a muscular and movable snout, is round or a vertical slit
and opens into a buccal cavity and hence into a muscular pharynx. The buccal cavity may be
armed with a pair of jaws (hardened plates made of hardened rods or prisms covered by
cuticle and operated by muscles). The buccal cavity or pharynx receive ducts from one or two
pairs of salivary glands. These buccopharyngeal slaivary glands secrete enzymes to
digest protein and carbohydrates. Usually a circular lip or a pair of vertical lips border the
mouth. Inner lips, sometimes tentacle-like (labial palps) may also be present. In some forms
the mouth with its circular lip may act as a sucker. Marine carnivorous forms have the mouth
at the end of a long proboscis which may invaginate (turn inside-out) upon retraction. A
radula may be present, projecting into the pharynx floor. The radula bears from a few to
several thousand chitinous teeth. In some carnivorous snails, the radular apparatus is
modified into a venom-injecting mechanism. The radula is rooted in the ventral wall of the
pharynx and is operated by powerful muscles, some of which insert into cartilaginous bolsters
of chondroid tissue. These muscles can retract or extend the radula. The radula itself is
secreted by the radular sac and is continually replaced.
The pharynx opens into the oesophagus (which may have a pair of glandular pouches) and
hence into the stomach. The oesophagus may have a pair of longitudinal ridges with a
ciliated gutter or food groove to convey the food. The stomach may be lined by cuticle and
has two longitudinal ridges or typhlosoles with a ciliated furrow in-between which conveys
food to the intestine. The stomach is not muscular, but relies on cilia to transport food via a
complex array of ciliated folds and grooves. The intestine winds back and forth within the
visceral mass. It is lined by mucus-secreting cells and may open into an expansion or rectum
which opens to the outside through the anus into the roof of the mantle cavity.
The anterior chamber of the stomach may have a style sac, which may communicate with the
intestine. This may contain a rod, the crystalline style (present in some herbivorous forms)
the lower end of which projects into the stomach and abuts onto a cuticularised area of the
stomach wall called the gastric shield. The style-rod may be up to 22 cm long and is made
of a transparent, flexible protein. It is rotated by cilia in the style sac and advanced into the
stomach where it is constantly worn away against the gastric shield and continually replaced
by secretion from the style sac. It helps draw the mucus-bound food-string into the stomach,
churns the stomach contents. The style is absent in carnivores. Amoebocytes beneath the
epithelium of the gut ingest food particles, though there may be extracellular digestion in the
stomach.
The mouth, which is often borne on a muscular and movable snout, is round or a vertical slit
and opens into a buccal cavity and hence into a muscular pharynx. The buccal cavity may be
armed with a pair of jaws (hardened plates made of hardened rods or prisms covered by
cuticle and operated by muscles). The buccal cavity or pharynx receive ducts from one or two
pairs of salivary glands. These buccopharyngeal slaivary glands secrete enzymes to
digest protein and carbohydrates. Usually a circular lip or a pair of vertical lips border the
mouth. Inner lips, sometimes tentacle-like (labial palps) may also be present. In some forms
the mouth with its circular lip may act as a sucker. Marine carnivorous forms have the mouth
at the end of a long proboscis which may invaginate (turn inside-out) upon retraction. A
radula may be present, projecting into the pharynx floor. The radula bears from a few to
several thousand chitinous teeth. In some carnivorous snails, the radular apparatus is
modified into a venom-injecting mechanism. The radula is rooted in the ventral wall of the
pharynx and is operated by powerful muscles, some of which insert into cartilaginous bolsters
of chondroid tissue. These muscles can retract or extend the radula. The radula itself is
secreted by the radular sac and is continually replaced.
The pharynx opens into the oesophagus (which may have a pair of glandular pouches) and
hence into the stomach. The oesophagus may have a pair of longitudinal ridges with a
ciliated gutter or food groove to convey the food. The stomach may be lined by cuticle and
has two longitudinal ridges or typhlosoles with a ciliated furrow in-between which conveys
food to the intestine. The stomach is not muscular, but relies on cilia to transport food via a
complex array of ciliated folds and grooves. The intestine winds back and forth within the
visceral mass. It is lined by mucus-secreting cells and may open into an expansion or rectum
which opens to the outside through the anus into the roof of the mantle cavity.
The anterior chamber of the stomach may have a style sac, which may communicate with the
intestine. This may contain a rod, the crystalline style (present in some herbivorous forms)
the lower end of which projects into the stomach and abuts onto a cuticularised area of the
stomach wall called the gastric shield. The style-rod may be up to 22 cm long and is made
of a transparent, flexible protein. It is rotated by cilia in the style sac and advanced into the
stomach where it is constantly worn away against the gastric shield and continually replaced
by secretion from the style sac. It helps draw the mucus-bound food-string into the stomach,
churns the stomach contents. The style is absent in carnivores. Amoebocytes beneath the
epithelium of the gut ingest food particles, though there may be extracellular digestion in the
stomach.
Above:
when removed from its shell, a typical gastropod mollusc is
somewhat like a bizarre
worm in appearance! The mantle lines a space within the shell and beneath it is the mantle
cavity. In life sea water is pumped through this cavity to irrigate the gills and carry away waste
materials.
Musculature
The columellar muscle (spindle muscle, shell muscle) extends from the columella to the
right side of the body and fan into the foot and attach to the operculum (or basement
membrane of the sole epithelium if the operculum is absent). The shape and divisions of this
muscle are particular to each form and it may be reduced or absent in some forms. A smaller
left columellar muscle may also be present. This muscle clearly acts to retract the animal.
Layers of muscle (an outer circular muscle layer and an inner longitudinal muscle layer) may
underlie this epidermis in the mantle and other exposed areas. Together with fibres from the
columellar muscle, these retract the mantle edge. The foot also contains transverse fibres
(crossing from one side to the other) to narrow the foot, dorsal longitudinal fibres (running the
length of the foot) to shorten it and oblique muscles and sometimes circular muscles. In sea
butterflies (heteropods) muscle fibres extend into the fins.
worm in appearance! The mantle lines a space within the shell and beneath it is the mantle
cavity. In life sea water is pumped through this cavity to irrigate the gills and carry away waste
materials.
Musculature
The columellar muscle (spindle muscle, shell muscle) extends from the columella to the
right side of the body and fan into the foot and attach to the operculum (or basement
membrane of the sole epithelium if the operculum is absent). The shape and divisions of this
muscle are particular to each form and it may be reduced or absent in some forms. A smaller
left columellar muscle may also be present. This muscle clearly acts to retract the animal.
Layers of muscle (an outer circular muscle layer and an inner longitudinal muscle layer) may
underlie this epidermis in the mantle and other exposed areas. Together with fibres from the
columellar muscle, these retract the mantle edge. The foot also contains transverse fibres
(crossing from one side to the other) to narrow the foot, dorsal longitudinal fibres (running the
length of the foot) to shorten it and oblique muscles and sometimes circular muscles. In sea
butterflies (heteropods) muscle fibres extend into the fins.


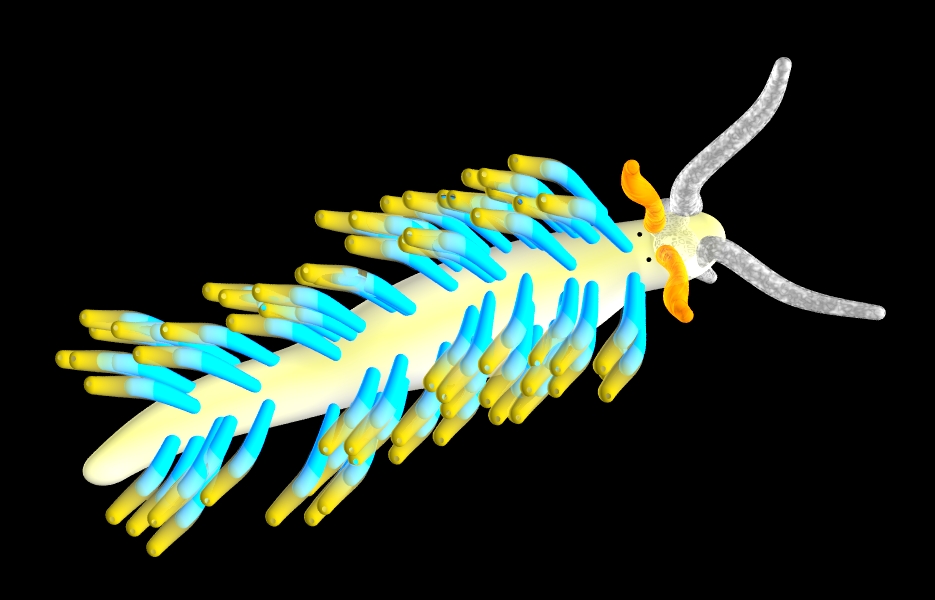
Above
and left: eolidacean nudibranchs
with cerrata. 'Nudibranch' literally means
naked gill, meaning that the gill is entirely
external rather than being housed in the
mantle cavity.
Some nudibranchs have tubular or
paddle-like extensions of their bodies,
typically forming two rows of clusters along
the animal's back. These cerata possibly act
as gills. each ceras may have a ciliated
epidermis over its surface (assisting
circulation of sea water around the ceras)
and contain large blood spaces. An
extension of the midgut gland enters the
ceras, and in eolidacean nudibranchs, this
may terminate in a sac, called a cnidosac,
which usually opens via a terminal pore and
which contains stinging nematocysts
obtained from ingested cnidarians. These
clearly also serve a defensive function.
Many nudibranchs possess instead a circle
of feathery gills around the anus
(circumanal gills).
The frontal pair of tentacles are the cephalic
tentacles (though in the model above,
expanded labial tentacles underneath can
just be seen) and behind these are the
rhinophores.
Some nudibranchs may lack gills and
tentacles and are superficially hard to
distinguish from planarian flatworms.
with cerrata. 'Nudibranch' literally means
naked gill, meaning that the gill is entirely
external rather than being housed in the
mantle cavity.
Some nudibranchs have tubular or
paddle-like extensions of their bodies,
typically forming two rows of clusters along
the animal's back. These cerata possibly act
as gills. each ceras may have a ciliated
epidermis over its surface (assisting
circulation of sea water around the ceras)
and contain large blood spaces. An
extension of the midgut gland enters the
ceras, and in eolidacean nudibranchs, this
may terminate in a sac, called a cnidosac,
which usually opens via a terminal pore and
which contains stinging nematocysts
obtained from ingested cnidarians. These
clearly also serve a defensive function.
Many nudibranchs possess instead a circle
of feathery gills around the anus
(circumanal gills).
The frontal pair of tentacles are the cephalic
tentacles (though in the model above,
expanded labial tentacles underneath can
just be seen) and behind these are the
rhinophores.
Some nudibranchs may lack gills and
tentacles and are superficially hard to
distinguish from planarian flatworms.
Sea
hares,
such as Aplysia, may reach 40 cm in
length and have a reduced shell, consisting of a
single flattened piece concealed by overgrowth of the mantle. Sea hares can discharge a purple fluid
to conceal them from enemies, discharged via the hole through which the mantle cavity opens to the
exterior. Others secrete a milky fluid with a nauseating odour, again in self-defense. Some
opisthobranchs have the typical gastropod arrangement, with a spiral shell enclosing the mantle cavity
and visceral mass. The pteropods are free-swimming marine snails and sea slugs. Some have a
spiral, tusklike, fanlike, or triangular shell which may bear spikes and can enclose the whole animal. In
some prosobranchs, the shell can not accommodate the whole animal which cannot fully retract into it.
Shell-less pteropods, nudibranchs and some other forms have no shell at all in the adult, although a
shell may be present in larvae or juvenile forms. Some sea slugs contain calcareous spicules in their
tissues.
The head may or may not be distinctly demarcated from the main body. The mouth may be terminal or
ventral and possess one or two lips. A proboscis may be present, which may or may not be
invaginable. There is usually one pair of eyes which are rarely borne on tentacles. Usually 1 to 3 pairs
of tentacles occur: the labial tentacles may be expanded into oral lappets (forming a labial veil) or form
a plough for burrowing as in Aplysia. The cephalic tentacles occur just behind the oral region. The
most posterior pair of tentacles are the rhinophores. The tentacles are often rolled up to form a fold.
Some forms have a frontal sail - a dorsal erect transverse fold on the front of the head.
single flattened piece concealed by overgrowth of the mantle. Sea hares can discharge a purple fluid
to conceal them from enemies, discharged via the hole through which the mantle cavity opens to the
exterior. Others secrete a milky fluid with a nauseating odour, again in self-defense. Some
opisthobranchs have the typical gastropod arrangement, with a spiral shell enclosing the mantle cavity
and visceral mass. The pteropods are free-swimming marine snails and sea slugs. Some have a
spiral, tusklike, fanlike, or triangular shell which may bear spikes and can enclose the whole animal. In
some prosobranchs, the shell can not accommodate the whole animal which cannot fully retract into it.
Shell-less pteropods, nudibranchs and some other forms have no shell at all in the adult, although a
shell may be present in larvae or juvenile forms. Some sea slugs contain calcareous spicules in their
tissues.
The head may or may not be distinctly demarcated from the main body. The mouth may be terminal or
ventral and possess one or two lips. A proboscis may be present, which may or may not be
invaginable. There is usually one pair of eyes which are rarely borne on tentacles. Usually 1 to 3 pairs
of tentacles occur: the labial tentacles may be expanded into oral lappets (forming a labial veil) or form
a plough for burrowing as in Aplysia. The cephalic tentacles occur just behind the oral region. The
most posterior pair of tentacles are the rhinophores. The tentacles are often rolled up to form a fold.
Some forms have a frontal sail - a dorsal erect transverse fold on the front of the head.
Article
last updated: 10/5/2014
More
Molluscs:
Pteropods
Cephalopods
Bivalves